Predictive Value of Biomarkers for Normal and Pathological Ageing in a Rat Model
Stramek A1, Wassif W2, Evans K1, Barkans J1, Mac Queen HA1*
1 Department of Life, Health and Chemical Sciences, The Open University, Walton Hall, Milton Keynes, UK.
2 Department of Clinical Biochemistry and Metabolic Medicine, Bedford Hospital NHS Trust, Kempston Road, Bedford, UK.
*Corresponding Author
H.MacQueen,
Professor, Department of Life,
Health and Chemical Sciences, The Open University,
Walton Hall, Milton Keynes, MK7 6AA, UK.
Tel: (44) 1908 655071
Fax: (44) 1908 654167
E-mail: Hilary.MacQueen@open.ac.uk
Received: September 24, 2015; Accepted: December 14, 2015; Published: December 19, 2015
Citation: Stramek A, Wassif W, Evans K, Barkans J, MacQueen HA (2015) Predictive Value of Biomarkers for Normal and Pathological Ageing in a Rat Model. Int J Food Sci Nutr Diet. S3:003, 16-22. doi: dx.doi.org/10.19070/2326-3350-SI03003
Copyright: MacQueen HA© 2015. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
Sub-optimal diets are widely known to be associated with disease. This study investigated two questions: first, whether a Western affluent diet affects long-term health in the rat, and second, whether, and to what extent, early changes in blood biomarkers can predict specific pathologies. A rat model was used in which animals were fed from weaning either a control diet or a high fat, low protein, energy dense Western affluent diet. Rats were harvested at either 12 or 18 months, and at these time points blood samples were taken and various clinical biomarkers measured using a hospital analyser or by ELISA. Predictive biomarkers should show small changes at early time points and larger changes at later time-points, and be associated with disease(s). Tissues were examined for overt pathology, and data were mined to establish links between pathologies and specific blood markers that might be used predictively. Results showed that the Western affluent diet is associated with ill-health, with a relative risk of developing disease 4.5 times higher than for the control diet. Furthermore we conclude that triacylglycerol, HDL cholesterol, the ratio of total cholesterol to HDL cholesterol, creatinine and alanine aminotransferase were good predictors of disease as across the cohorts they showed altered levels before the clinical development of pathologies, in agreement with our hypothesis.
2.Introduction
3.Materials and Methods
3.1 Animals
3.2 Diets
3.3 Sampling
3.4 Biochemical analyses
3.5 Statistical analyses
4.Results and Discussion
4.1 Animal health
4.2 Biochemical analysis
5.Conclusion
6.Acknowledgements
7.References
Keywords
Biomarkers; Diet; ageing; Rat; Insulin.
Introduction
The ageing process in mammals is complex and is often accompanied by a number of chronic and acute pathologies whose incidence increases with age. Moreover life events can impact substantially on both the ageing process itself and on the incidence and impact to the individual of the medical conditions, and a number of conditions can be influenced by lifestyle factors such as diet and exercise. Sub-optimal diet has been linked to a large proportion of the global burden of disease [1]. If clinical conditions can be alleviated by non-pharmacological interventions such as a change in diet, as is the case for Type II diabetes [2-4], a significant cost saving could be made in sustaining an ageing population.
Over the course of evolution mechanisms have developed that allow individuals to withstand periods of nutritional deficiency, and this has been discussed elsewhere [5]. However, in developed countries in the present day the more common problem is one of nutrient over-availability, and changed patterns of food consumption have led to an obesity epidemic in the human population and an increase in the incidence of conditions such as diabetes, coronary heart disease and joint disease, all of which impact significantly on quality of life. Such diseases are no longer thought of as diseases of old age, since they are increasingly prevalent in much younger age groups. We and others have studied the effects of diet on markers of disease [6-9]. Rats are frequently used in such research because they bring all the benefits of a well-understood laboratory model, and are not fussy eaters. Rats are also subject to many deleterious effects of ageing, and long-term studies using rats reveal the gradual onset of age-related pathologies.
The experimental diet that we have used here models the kind of sub-optimal diet that is increasingly eaten in affluent countries: it is high in fat and low in protein and fibre compared to an optimal control diet. The two diets used here are equivalent in carbohydrate content, but the high fat content of the Western affluent diet makes it more energy-dense than the control diet, which is a chow made by the same manufacturer but optimised for rodent maintenance [10]. In addition to cardiovascular problems [11], high-fat diets of this kind are known to promote the development of fatty liver disease and other metabolic abnormalities [8], to affect the structure of the kidney [12], and to affect the performance of the immune system [13]. All of these parameters are important for the development of chronic health conditions, and at a population level there is interest in identifying specific biomarkers that can be used to predict the onset of cardiovascular, hepatic and renal pathologies, and hence to manage the risks associated with them.
Here we describe a study undertaken to investigate two questions: first, whether a Western affluent diet affects long-term health in the rat, and second, to what extent early changes in blood biomarkers can predict specific pathologies. This information could be used to predict future illness so that dietary or other interventions may be made before the clinical onset of the disease. In order to be a reliable predictor a biomarker must be linked to a known abnormality, and must be detectable above background level at a time before the abnormality has become a clinical problem.
In this study we examined the gross morphology of target organs, and levels of biochemical markers used clinically for the diagnosis of cardiovascular, liver and kidney abnormalities. To address our first question we compared the prevalence of pathologies in a control population eating an optimal diet with that in a population eating a high fat, low protein Western affluent diet. To address the second question we examined the profile of blood biomarkers in these two groups at three time points: weaning, 12 months and 18 months of age. We sought biomarkers that showed small changes at the 12 month time point and larger changes at the 18 month time point, and that could be linked to an increased level of particular pathologies.
Materials and Methods
Animals were male Sprague-Dawley rats, bred in-house and maintained in an enriched environment on a 14h light: 10h dark cycle. Animals were kept and treated in accordance with the UK Animals (Scientific Procedures) Act, 1986, and the experiments were approved by the Open University Animal Welfare and Ethics Review Board. Rats were caged in Scantainers (Scanbur Technology, Denmark) at ambient temperature (19-23°C) and 50 ± 10% humidity. They were given cardboard tubes and aspen wood blocks (LBS Biotechnology, UK). Rats were weaned at three weeks old directly onto the experimental or control diets, which were available ad libitum, and were maintained at two littermates per cage until the age of 12 months or 18 months, when they were used.
During this period each rat was weighed weekly until 5 months of age and monthly thereafter, and examined visually for signs of ill-health. Because the rats were given wood blocks to gnaw, no dental problems were encountered. Growth curves for the animals are shown in Figure 1a. Hepatomegaly is frequently associated with high-fat diets [14], but Figure 1b shows that in this study the size of the liver remained in proportion to body mass across all the animal groups. All animals were harvested at the same time of day, 5 hours into the light phase. A total of 35 rats were used in this study. Seven weanlings were used as controls; these animals were assumed to be in optimal dietary conditions although not necessarily representative of older, weaned animals eating an optimum diet. At each time point (12 months and 18 months) 4 animals eating the control diet and 10 animals eating the experimental diet were harvested. The reason for the smaller number of control animals is that we already have substantial data on such animals [8, 14] and for ethical reasons did not wish to repeat this data collection needlessly.
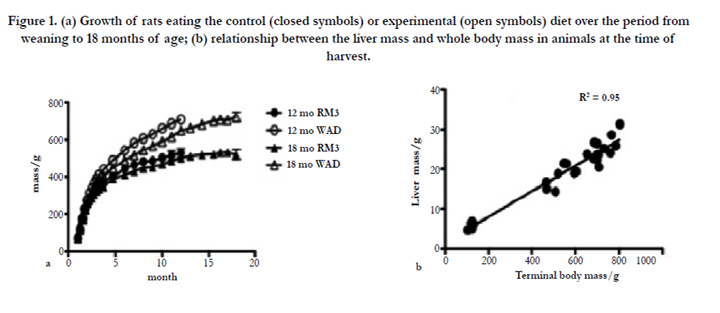
Figure 1. (a) Growth of rats eating the control (closed symbols) or experimental (open symbols) diet over the period from weaning to 18 months of age; (b) relationship between the liver mass and whole body mass in animals at the time of harvest.
The control diet was RM3 and the experimental diet was Western RD (WRD), both purchased from Special Diet Services, Witham, Essex, UK. The components of both diets, taken from the manufacturer’s data [10], are shown in Table 1. It can be seen that the principal differences between the diets are the content of fat (higher in WRD) and protein (lower in WRD). WRD is also lower in fibre and ash, suggesting that this diet may be less able to support gut function. The carbohydrate components of the two diets are equivalent, but the WRD is more energy-dense, as estimated from the Atwater Fuel Energy calculation [15].
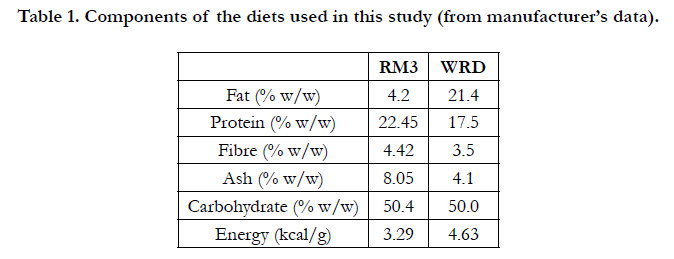

Table 1. Components of the diets used in this study (from manufacturer’s data).
At the time of harvest, rats were weighed then deeply anaesthetised with Isofane (Novartis Animal Health UK Ltd). Approximately 4 ml of blood was obtained by cardiac puncture. Following coagulation at room temperature the clots were spun down and the supernatants were removed and stored at –80°C until analysis. Immediately following cardiac bleeding, the rats were killed by intracardiac injection of Pentoject (Animal Care Ltd, UK). Organs were then removed for visual assessment of size, shape, colour and the presence of any gross abnormalities such as ulcers or tumours. They were not routinely assessed histologically. Organs removed for examination were brain, heart, stomach, intestines, liver, pancreas, spleen and kidneys.
Serum metabolites, electrolytes, renal profile, liver profile and lipid profile were measured using a Cobas Integra 800 analyser (Roche Diagnostics, Mannheim, Germany). The analytes tested are listed in Table 2. Sera from the same animals were also used in triplicate in ELISAs to determine circulating levels of insulin. All ELISA assays were carried out using the Alpco Insulin (rat) ultrasensitive ELISA kit (Stratech Scientific Ltd, Newmarket, UK).
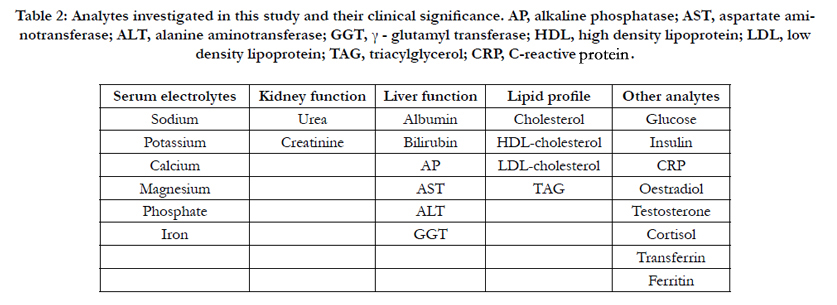
Table 2: Analytes investigated in this study and their clinical significance. AP, alkaline phosphatase; AST, aspartate aminotransferase; ALT, alanine aminotransferase; GGT, γ - glutamyl transferase; HDL, high density lipoprotein; LDL, low density lipoprotein; TAG, triacylglycerol; CRP, C-reactive protein.
Data are expressed as mean ± SEM. Differences between means were assessed by ANOVAs and t-tests as appropriate, using Prism v5 (GraphPad Software, San Diego, California, USA, www.graphpad.com). Since sample sizes were small and unequal, and had unequal variances (with the variances being greater in the larger experimental diet groups) ANOVA cannot detect small but significant differences, and it is recommended to use t-tests for analysing these data [16]. Differences were considered statistically significant at the P < 0.05 level. Relative risk was calculated from the equation:
Relative risk = probability of disease in rats eating WRD/probability of disease in rats eating RM3.
Results and Discussion
Rats are commonly used in dietary studies because they are not fussy eaters and are physiologically robust [17]. In this study we observed the health status of rats eating one of two diets, and at three time points. We measured the levels of standard biochemical markers used diagnostically, and related them to the pathologies that we observed. We were particularly looking at a population level for biochemical markers that showed significant changes by 18 months on a sub-optimal diet and which also showed a lesser change in the same direction at 12 months on the diet, such that the marker could be used clinically in a predictive way to allow early therapeutic interventions.
A number of gross pathologies were observed when the animals were harvested. At the start of the experiment all rats were healthy and active, with good coats and general appearance. One rat eating the control diet and one eating the WRD had to be culled before 12 months due to poor health. Two further rats eating the experimental WRD were found dead in their cages between 12 and 18 months. These four animals were omitted from the analysis. Among the animals which survived until the experimental end-point, 4 out of 13 rats harvested at 12 months, and 7 out of 11 rats harvested at 18 months were found to have one or more pathologies upon dissection in spite of appearing in good health judging by coat condition, food intake and activity. Overall, 12.5% of animals eating the RM3 diet (all ages) had an overt pathology, compared with 56.3% of those eating the WRD (33.3% at 12 months and 85.7% at 18 months; see Table 3). This means that over the course of the study the relative risk of developing a pathology was 4.5 times higher for rats eating the WRD than for those eating the RM3 diet.
Table 3 also shows that all but one of the animals exhibiting pathologies had levels of specific biomarkers either elevated or depressed beyond one standard deviation from the population mean. The specific analyte changes were consistent with the observed pathologies; for example kidney abnormalities were accompanied by changes in creatinine and urea levels in the blood. This indicates that the analytes used do indeed reflect particular health problems. These results clearly demonstrate that rats can appear to be healthy while nevertheless harbouring a variety of pathologies, and provide further evidence that a sub-optimal Western affluent diet cannot support long-term health.
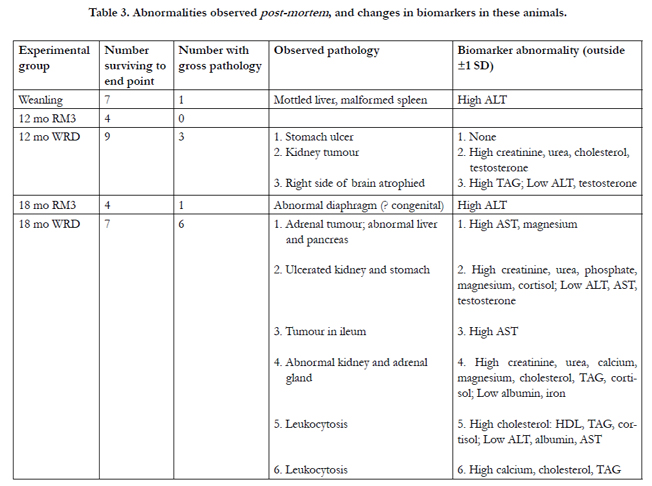
Table 3. Abnormalities observed post-mortem, and changes in biomarkers in these animals.
Following biochemical analysis, the majority of analytes showed no differences between the cohorts of animals eating the two diets or at the ages examined. These analytes included C-reactive protein, a marker of immune function, so in this study we were unable to detect any changes in the immune status of the animals. Statistically significant changes were, however, observed in the following analytes: iron, ALT, AST, albumin, creatinine, cholesterol, HDL-cholesterol, TAG, cortisol, glucose and insulin. We detail these differences below.
Compared with the level seen at weaning, iron levels decreased with age, and were significantly lower (P < 0.01) in 18 month old animals compared with weanlings (Figure 2a). There were no significant differences between the diets. This was the only change observed in serum electrolytes, in contrast to previous work that showed a reduction in potassium level with age [8], and this may be accounted for by the different diets used in these two studies.
Corticosterone, not cortisol (which is measured by the clinical analyser) is the primary glucocorticoid stress hormone in the rat; nevertheless cortisol measurements can be used as a proxy measurement for stress as there is considerable overlap in the values for the two hormones [18]. There was a slight but not significant upward trend in cortisol in WRD animals compared to those eating the RM3 diet (Figure 2b). Levels of glucocorticoid stress hormones have been reported to rise with age in the rat [19] and we also observed this trend, although the rise was not statistically significant. However we found that cortisol levels were high in weanlings compared with the other groups (Figure 2b). Similar findings have been reported by others, and this effect is thought to be a response to the stress of weaning [20].
ALT rose with age (RM3 diet) but dropped in the WRD animals (Figure 2c). This pattern was also seen to a lesser extent with AST (not shown). The significant drop in 18 month WRD animals (P < 0.001) was accompanied by a non-significant but consistent decrease at 12 months; thus ALT is a possible candidate for a disease biomarker, although it may be an imprecise marker since low ALT was associated with an atrophied brain, ulcerated kidney and stomach, and leukocytosis (Table 3). Albumin remained stable in the RM3 animals, but there was a highly significant drop (P < 0.001) in the WRD animals at 18 months which was not seen at 12 months (Figure 2d). Liver enzymes are frequently used as clinical markers of a range of conditions, and we have previously found a number of changes with age and diet in Sprague-Dawley rats [8, 14]. Although in humans an enlarged liver is frequently associated with ill-health, we did not observe this phenomenon, and all the livers remained in the correct proportion to whole body mass (Figure 1b). A histological examination of some of the livers (not shown) showed glycogen and fatty deposits increasing with age, and noticeably more prominent with animals eating the high-fat diet, consistent with our previous findings and those of others [14, 21]. In this study we observed that the rise in AST and ALT with age seen in control animals was substantially abolished in WRD animals, which also showed a significant drop in levels of albumin at 18 months (Figure 2c and d; P < 0.001). These results suggest a profound change in the liver metabolism of WRD animals that can be linked to general ill-health (as revealed by the pathology data) but not to any specific condition.
In view of the number of kidney abnormalities that we observed post-mortem we anticipated that there would be changes in the levels of the kidney markers that we used, particularly since levels of these markers are known to vary with the protein content of the diet [22]. Across the experimental groups there were no changes in levels of urea (Figure 2e). Mean creatinine levels did not change with age in the RM3 animals, but increased with age in the WRD animals (Figure 2f). Individual rats with kidney abnormalities had elevated creatinine levels compared with normal members of their cohort (Table 3), accounting for the significant rise in the mean value (and SEM) seen in the 18 month WRD group. This suggests that, in addition to its current diagnostic use [3], creatinine could be used in a predictive way to allow early therapeutic intervention for kidney abnormalities. Based on these data, further study of creatinine expression during disease development is warranted.
Blood lipids are well known as markers of illhealth, and are particularly linked to cardiovascular disease. In these experiments total cholesterol increased in the control animals between weaning and 12 months of age (P < 0.05), but remained stable thereafter. The WRD 12 month group had a similar cholesterol level to the RM3 group, but at 18 months the mean had increased to a highly significant level (P < 0.001; Figure 2g). In these studies LDL cholesterol remained fairly stable (not shown) but the HDL cholesterol showed an interesting pattern. In the control RM3 rats HDL cholesterol increased significantly by 12 months, but this increase was completely abolished in the WRD animals at both 12 and 18 months (Figure 2h). This was not unexpected, given the fat content of the WRD (Table 1). The consequence of this change was that the total cholesterol: HDL-cholesterol ratio showed significant changes with diet, with the 12 month value being a good predictor of the 18 month value (Figure 2i). Likewise, whereas the control animals showed a non-significant increase in TAG level at 12 months compared to weanlings that then remained stable, the WRD animals showed a significant rise (P < 0.01) at 18 months that was predicted by the 12 month pattern (Figure 2j). A separately conducted multivariate analysis of volatile organic compounds in the faeces of the same animals [23] also suggested that TAG, total cholesterol, and the total cholesterol : HDL-cholesterol ratio could be reliably used as predictive markers.
As noted above, the animals in this study were fed ad libitum, and all were harvested at the same time in the dark-light cycle. The carbohydrate content of the two diets was similar (Table 1), so in spite of the raised body mass and increased blood lipid markers exhibited by the WRD animals (Figures 1a and 2g-j) there was no increase in circulating glucose levels. In fact by 18 months in both control and experimental animals blood glucose levels were significantly lower (P < 0.001) than at weaning and 12 months (Figure 3a).
An unexpected result was found when we examined the levels of circulating insulin. It has been shown in several species including the rat that circulating insulin levels rise with increasing glucose intake [24] and insulin levels are linked to diabetes. We observed no rise in insulin levels with either age or diet, and given the carbohydrate content of the two diets noted above this was not surprising. However, insulin values fell over a wide range in all the cohorts, and the spread of values seen was similar regardless of age or diet (Figure 3b). Careful examination of the data revealed that no values fell between 0.7 and 0.9 ng/ml, suggesting that there were two insulin-secreting phenotypes: one giving values in the range 0.2 to 0.7 ng/ml (mean 0.35 ng/ml), and the other giving values in the range 0.9 to 2.3 ng/ml (mean 1.4 ng/ ml). These populations were significantly different by t-test (P < 0.001; Figure 3c). Differences in insulin levels were not associated with changes in glucose levels (Figure 3d), nor was there any correlation between insulin level and the level of any lipid markers (Figure 3e), terminal body mass (Figure 3f), or any specific gross pathology (Table 3). We consider it unlikely that the high values represent a post-prandial increase since rats are trickle feeders and do not exhibit the pattern of insulin secretion found in humans. The mean value exhibited by the higher population (1.4 ng/ml) is similar though slightly higher than that reported for baseline circulating insulin by others: 1 ng/ ml [24] or 1.09 ng/ml [25]. The mean lower value (0.35 ng/ml) is slightly below the range of 0.4- 0.8 ng/ml reported for rats starved for 24 hours [25], but it should be noted that the rats in our study had not been starved and may just naturally exhibit a lower baseline circulating level. As far as we are aware this is the first report of different insulin secreting phenotypes in unstressed Sprague-Dawley rats.
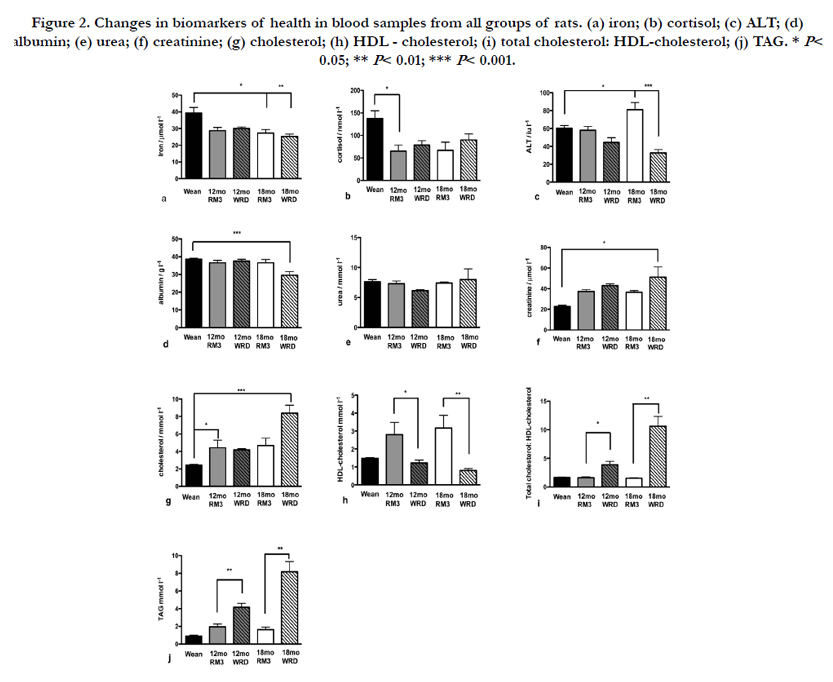
Figure 2. Changes in biomarkers of health in blood samples from all groups of rats. (a) iron; (b) cortisol; (c) ALT; (d) albumin; (e) urea; (f) creatinine; (g) cholesterol; (h) HDL - cholesterol; (i) total cholesterol: HDL-cholesterol; (j) TAG. * P< 0.05; ** P< 0.01; *** P< 0.001.
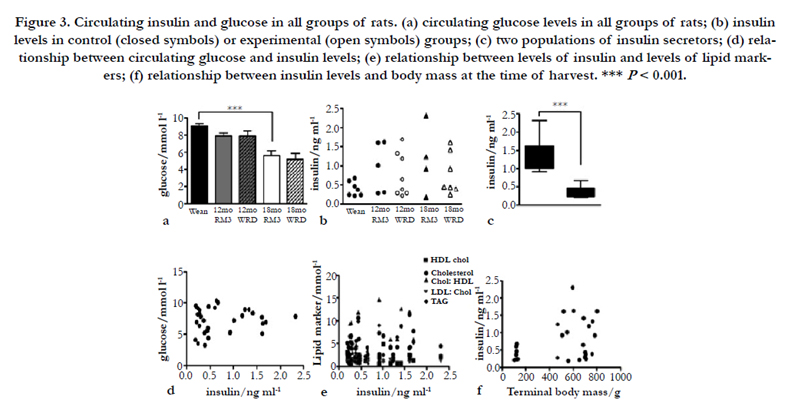

Figure 3. Circulating insulin and glucose in all groups of rats. (a) circulating glucose levels in all groups of rats; (b) insulin levels in control (closed symbols) or experimental (open symbols) groups; (c) two populations of insulin secretors; (d) relationship between circulating glucose and insulin levels; (e) relationship between levels of insulin and levels of lipid markers; (f) relationship between insulin levels and body mass at the time of harvest. *** P < 0.001.
Conclusion
This study focused on two questions: first, whether a Western affluent diet affects long-term health in the rat, and second, whether early changes in blood biomarkers can predict specific pathologies. The first question has an unequivocal answer, in that the health of animals eating this diet was notably worse than that of animals eating a control, optimised diet, as measured by gross pathologies observed post-mortem. The relative risk was 4.5 over the course of the study.
Regarding the second question, we have found limited feasibility for using known biomarkers as predictors of future disease, at least in the Sprague-Dawley rat. Five markers: HDL cholesterol, total cholesterol: HDL cholesterol (driven by the change in HDL cholesterol), triacylglycerols, ALT and creatinine clearly show a suitable pattern of change. The changes for these markers are consistent and statistically significant, although it is unclear at present whether they correlate with specific pathologies, and more work is required to confirm this. Our data bring together studies of individual animals in which biomarker analysis can be correlated with specific health status, and cohort studies in which trends can be more easily identified. Future work in this area should use longitudinal studies with repeated sampling from the same animals.
We also report a previously unrecognised phenotypic diversity in insulin production by Sprague-Dawley rats.
Acknowledgements
The authors declare no conflict of interest. We would like to thank Dr Jane Loughlin for helpful comments on the manuscript. This research received no specific grant from any funding agency in the public, commercial or not-for-profit sectors. We acknowledge The Open University Biomedical Research Network for pumppriming funding.
References
- Forouzanfar M, Alexander L, Anderson H, Bachman V, Biryukov S, et al.(2015) Global, regional, and national comparative risk assessment of 79 behavioural, environmental and occupational, and metabolic risks or clusters of risks in 188 countries, 1990-2013: a systematic analysis for the Global Burden of Disease Study 2013. Lancet 386(10010): 2287-2323.
- Turner R, Cull C, Frighi V, Holman R (1999) Glycemic control with diet, sulfonylurea, metformin, or insulin in patients with type 2 diabetes mellitus.JAMA 281(21): 2005-2012.
- Gross J, de Azevedo M, Silveiro S, Canani L, Caramori M, et al. (2005) Diabetic nephropathy: diagnosis, prevention and treatment. Diabetes Care 28(1): 164-176.
- Jibani M, Bloodworth L, Foden E, Griffiths K, Galpin O (1991) Predominantly vegetarian diets in patients with incipient and early clinical diabetic nephropathy: effects on albumin excretion rate and nutritional status. Diabet Med 8(10): 949-953.
- Cordain L, Eaton S, Sebastian A, Mann N, Lindeberg S, et al. (2005) Origins and evolution of the Western diet: health implications for the 21st century. Am J Clin Nutr 81(2): 341-354.
- Meydani S, Wu D (2007) Age-associated inflammatory changes: role of nutritional intervention. Nutr Rev 65(12 Pt 2): S213-S216.
- Ridker P (2007) Inflammatory biomarkers and risks of myocardial infarction, stroke, diabetes and total mortality: implications for longevity. Nutr Rev 65(12 Pt 2): S253-S259.
- MacQueen H, Wassif W, Walker I, Sadler D, Evans K (2011) Age-related biomarkers can be modulated by diet in the rat. Food and Nutrition Sciences 2(8): 884-890.
- He K, Liu K, Daviglus M, Jenny N, Mayer-Davis E, et al. (2009) Associations of dietary long-chain n-3 polyunsaturated fatty acids and fish with biomarkers of inflammation and endothelial activation (from the Multi-Ethnic Study of Atherosclerosis [MESA]). Am J Cardiol 103(9): 1238-1243.
- Special Diets Services Data Sheets. www.sdsdiets.com/pdfs/RM3-P.pdf
- Van Gaal L, Mertens I, de Block C (2006) Mechanisms linking obesity with cardiovascular disease. Nature 444: 875-880.
- Altunkaynak M, Ozbek E, Altunkaynak B, Can I, Unal D, et al. (2008) The effects of high-fat diet on the renal structure and morphometric parametric of kidneys in rats. J Anat 212(6): 845-852.
- MacQueen H, Sadler D, Mattacks C (2000) Dietary fatty acids influence the appearance of tumour necrosis factor-alpha receptors on adipocytes following an immune challenge. Br J Nutr 84(3): 387-392.
- MacQueen H, Sadler D, Moore S, Daya S, Brown J, et al. (2007) Deleterious effects of a cafeteria diet on the livers of non-obese rats. Nutr Res 27(1): 38-47.
- McCance R, Widdowson E (2014) The Composition of Foods. Royal Society of Chemistry. (7th edtn), Cambridge, London.
- De Winter J (2013) Using the Student’s t-test with extremely small sample sizes. Practical Assessment, Research and Evaluation 18(10): 1-12.
- Mattacks C, Sadler D, Pond C (2004) The effects of dietary lipids on dendritic cells in perinodal adipose tissue during chronic mild inflammation. Br J Nutr 91(6): 883-892.
- Koren L, Whiteside D, Fahlman A, Ruckstuhl K, Kutz S, et al. (2012) Cortisol and corticosterone independence in cortisol-dominant wildlife. Gen Comp Endocrinol 177(1): 113-119.
- Sapolsky R (1992) Do glucocorticoid concentrations rise with age in the rat? Neurobiol ageing 13(1): 171-174.
- Cooper T, Roberts M, Kattesh H, Kojima C (2009) Effects of transport stress, sex and weaning weight on postweaning performance in pigs. The Professional Animal Scientist 25: 189-194.
- Carmiel-Haggai M, Cederbaum AI, Neito N (2005) A high-fat diet leads to the progression of non-alcoholic fatty liver disease in obese rats. FASEB J 19(1): 136-138.
- Butani L, Polinsky M, Kaiser B, Baluarte H (2002) Dietary protein intake significantly affects the serum creatinine concentration. Kidney Int 61(5): 1907.
- Hopes K, Cauchi M, Walton C, MacQueen H, Wassif W, et al. (2015) A novel method for the analysis of clinical biomarkers to investigate the effect of diet on health in a rat model. Analyst 140(9): 3028-3038.
- Steffens A (1970) Plasma insulin content in relation to blood glucose level and meal pattern in the normal and hypothalamic hyperphagic rat. Physiol Behav 5(2): 147-151.
- Havel P, Uriu-Hare J, Liu T, Stanhope K, Stern J, et al. (1998) Marked and rapid decreases of circulating leptin in streptozotocin diabetic rats: reversal by insulin. Am J Physiol 274(5 Pt 2): R1482-R1491.





