Epidemiologic Dominance of HIV-1 Subtype CRF02_AG in Ghana: Preliminary Virological Evidence of Increasing Association With New Infections
Nii-Trebi NI1*, Barnor JS2, Musah BO3, Ampofo WK2
1 Department of Medical Laboratory Sciences, School of Biomedical and Allied Health Sciences, University of Ghana, Accra, Ghana.
2 Department of Virology, Noguchi Memorial Institute for Medical Research, University of Ghana, Accra, Ghana.
3 Department of Microbiology, School of Biomedical and Allied Health Sciences, University of Ghana, Accra, Ghana.
*Corresponding Author
Nicholas Israel Nii-Trebi
Department of Medical Laboratory Sciences, School of Biomedical and Allied Health Sciences,
University of Ghana. P.O. Box KB 143, Korle Bu, Accra, Ghana.
Tel +233 302 687 974/5
Fax +233 302 688 291
Email : nntrebi@chs.edu.gh
Received: February 29, 2016; Accepted: April 22, 2016; Published: April 27, 2016
Citation: Nii-Trebi NI, Barnor JS, Musah BO, Ampofo WK (2016) Epidemiologic dominance of HIV-1 subtype CRF02_AG in Ghana: preliminary virological evidence of increasing association with new infections . Int J Virol Stud Res. 4(2), 22-28.
doi: dx.doi.org/10.19070/2330-0027-160004
Copyright: Nii-Trebi NI© 2016. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
In Ghana, HIV-1 subtype prevalence has varied appreciably over the past two and half decades. Recombinant forms have also increasingly emerged over the past decade. The dynamic nature of HIV-1 species profile in Ghana deserves exploration. This preliminary study investigates virological properties of epidemiologically prevalent HIV-1 subtypes in Ghana. Viruses were isolated from PBMCs obtained from Ghanaian HIV-1-infected individuals. Viral supernatants were prepared by transfection with subtyped HIV-1 full length DNA clones derived from viral isolates. Levels of Virion production were quantitated by HIV-1 p24 antigen ELISA. MAGIC5A cells were infected by normalized virus and viral infectivity was determined by MAGI assay. virion productivities and viral infectivities were compared by the nonparametric Kuskal-wallis one way ANOVA test. Findings show successful transfection of HIV DNA clones into cells. Virion productivity was found to be comparable (p=0.405) among eight strains tested. MAGI assay showed the infectivity of HIV-1 circulating recombinant form, CRF02_AG to be significantly higher (p=0.044) as compared to pure subtype A and subtype G viruses. The apparently high infectivity of the recombinant form observed in this study may suggest that recombination events might have the advantage in acquiring higher infectivity and pathogenesis during viral evolution. This may explain the increasing association of the CRF02_AG subtype with new infections and also highlight the evolutionary potential of HIV-1 in Ghana. The study sheds light on the virological basis of the epidemiologic dominance of prevalent HIV-1 subtypes and the role of in vitro HIV-1 infectivity in the in vivo situation.
2.Introduction
3.Materials and Methods
3.1 Patients and samples
3.2 Cell maintenance, virus isolation and proviral quantitation
3.3 Virion productivity assay
3.4 Determination of virus concentration
3.5 Virion infectivity assay
3.6 Data analysis
3.7 Ethics
4.Results
4.1 Viral isolation
4.2 Virion Productivity determination by p24 Assay
4.3 Virion infectivity
5.Discussion
6.Acknowledgements
7.References
Keywords
HIV-1, Molecular epidemiology, Virion productivity, Viral infectivity
Introduction
The study and quantitation of virus infectivity has had important application for decades since cell culture replaced the use of embryonated eggs and laboratory animals for in vitro virus isolation and detection. In the absence of a suitable animal model to study the human immunodeficiency virus type 1 (HIV- 1), the usefulness of cell studies to obtain information about interactions between the host cell, the virus and its components cannot be overemphasized [1-3]. Viral infectivity studies thus plays a significant role in unraveling the kinetics of viral functions within the host, thereby providing useful information necessary for consideration in drug formulation.
As an obligate intracellular parasite, HIV must infect a susceptible, permissive cell to replicate its genome and ultimately produce viral particles (virions) that have the ability to infect other cells to ensure viral propagation in the host [4, 5]. However, often viral infection may be nonproductive or abortive. Besides, not all viruses produced in a productive infection cycle mature into infectious virions due to various viral and host factors [6, 7]. Furthermore, the error-prone nature of the reverse transcriptase tends to create nucleotide sequence variation of the HIV genome and this contributes to the emergence of viral variants. Thus exceptional genome variation of HIV stems from sources including high rates of mutation [8, 9], which in turn leads to divergent patterns in HIV evolution either within a population or within an individual host.
Characteristically, lentiviruses have the ability to recombine in host cells, which explain in part their genetic diversity. Occurrences of single mutations may not explain the emergence of recombinant strains. However, this has consequences in natural selection, disease progression as well as drug efficacy [10, 11]. Other factors including drug pressure also influence the viral genome variation and evolutionary potential, which might have effect on the viral infectivity, response to anti-retroviral therapy and also outcome of molecular detection of viral genome during infection.
Given the importance of HIV-1 genetic variation in the management of HIV-1 disease, the epidemiologic dominance of certain HIV-1 subtypes over time requires regular investigation. In Ghana, the HIV-1 species variation over the years has been quite dynamic, The virological basis and relevance for in vivo situation remains to be explored. This study therefore investigates the infectivity of HIV-1 subtypes in Ghana with the aim of shedding light on the role of in vitro infectivity in the in vivo significance of prevalent HIV-1subtypes.
Viruses used in this study were isolated from 15 consenting Ghanaian HIV-1-seropositive individuals accessing national antiretroviral treatment care at the Atua Government Hospital in the Eastern Region of Ghana. The HIV status of the patients was routinely determined by screening of blood samples for antibody to HIV using Abbott Determine HIV-1/2 (Abbott Diagnostics, Abbott Park, IL) rapid test and confirmation of HIV type by INNO-LIA HIV I/II (Innogenetics, Gent, Belgium). Blood samples were collected into EDTA tubes and peripheral blood mononuclear cells (PBMC) were separated from whole blood using Ficoll-Hypaque gradient (Sigma, St., MO). Cells were washed twice with phosphate-buffered saline (PBS) and PBMC pellets approximating to 5x106 were prepared and stored at -80°C until use.
Initially viruses were isolated directly by co-culturing PBMC with 1% phytohemagglutinin (PHA)-stimulated donor PBMC from an HIV seronegative individual and through MAGIC5A [CXCR4/CCR5-expressing MAGI (HeLa-CD4-LTR-β-gal) cells [12, 13]. To normalize DNA for transfection, proviral DNA was quantified in each sample through cell lysis followed by measurement of luciferase activities by the firefly Luciferase assay system (Promega, Madison, WI). Briefly, 48 hours after infection, cells were washed with Dulbecco’s Phosphate-Buffered Saline (DPBS) after removal of culture media. The cells were then lysed by addition of 100 ul of Glo lysis buffer (Promega) to each well and incubation for five minutes at room temperature. From each well, 50 ul lysate was transferred to a single well in a 96-well microtiter plate (Costar) and 50 ul of Bright-Glo™ assay reagent was added and well mixed. The luminescence was measured with Centro LB96V microplate luminometer (Berthold, Bad Wildbad, Germany) and the proviral DNA content was noted after subtraction of background luminescence (luminescence of TZM-bl cells without virus infection).
293T cells were used in virion productivity assay and MAGIC5A cells were used for virion infectivity assays. PBMCs were grown in RPMI (Gibco-BRL) and both cells for transfection and infectivity assays were maintained in Dulbecco's modified Eagle's medium (DMEM), each supplemented with 10% heat-inactivated fetal calf serum (FCS) (complete medium), 1% penicillin/ streptomycin, and amphotericin B (Gibco BRL).
To obtain viral supernatants, 5x105 293T cells were trnsfected with each proviral DNA in 25 cm2 flasks by using calcium phosphate method [14, 15]. Briefly, cells were split on the day prior to transfection so as to achieve 50-60% confluence. Five hundred microlitres of 250mM CaPO4 and 20μg HIV-1 DNA were added in sterile eppendorf tubes and mixed by flicking the tubes. The DNA/CaPO4 mix was added in drop-wise while vortexing to a solution of 500 υl 2XHBS (10g/l Hepes and 16 g/l NaCl; pH 7.1 + 0.05) and 20 υl 100XPO4 (1:1 of 70 mM Na2HPO4 and 70 mM NaH2PO4) in a 15 ml centrifuge tube and incubated for 30 min at room temperature. Two hundred microliters of the solution obtained were added drop-wise to cell culture media in a12-well plate and incubated. Sixteen hours after transfection the cells were washed twice with PBS and re-fed with 2 ml of fresh complete medium. The incubation was continued at 37°C in a humidified 5% CO2 incubator. Virions were harvested from culture supernatants 48 h post-transfection. Supernatants were filtered through 0.45μm pore filters (Millipore). Levels of virion production were quantitated (as described below) and the virus stocks were stored at - 80°C until use.
The concentration of virions produced was determined by the measurement of p24 values using HIV-1 p24-antigen capture enzyme-linked immunosorbent assay (ELISA) (Retro-Tek) following the manufacturer’s instructions. Briefly, virus samples were diluted, lysed and captured in a micro ELISA plate at 37°C for one hour. Subsequently wells were washed and human anti-p24 polyclonal antibodies conjugated with peroxidase were added to the captured p24 antigen and incubated at 37°C for one hour. Peroxidase substrate was added at the end of the incubation and then further incubated for 30 minutes at room temperature. The reaction was stopped by addition of 2N Sulfuric acid, after which the absorbance was measured at 450 nm in a microplate reader (BioTek instruments ). The concentration of the p24 antigen obtained for the various viral subtypes were adjusted (100pg) and used to infect subconfluent MAGIC5A cells in a virus infectivity assay.
HIV-1 virion infectivity was determined by the MAGI assay. A day prior to infection, MAGIC5A cells were seeded in 96-well plates (Falcon) in passage medium comprising DMEM supplemented with 2.5% FCS. On the next day, the medium was removed from each well. The cells (5 x 105) were then infected by adding viral supernatants containing 100 pg of p24 antigen in total volume of 25-50 μl of infection medium (passage medium supplemented with 20 μg/ml DEAE Dextran) and incubating for 2 hours at 37°C in a 5% CO2 incubator , with gentle rocking every 30 min. Fifty microlitres of infection medium were added to control wells.
After infection, the medium was removed from each well; 100 μl of passage medium were added and the cells were incubated for 48 hrs at 37°C in a 5% CO2 incubator. The medium was removed and cells were fixed for 5 min with 50 μl of fixation solution made up of 1% formaldehyde and 0.2% glutaraldehyde in PBS. The cells were then washed with PBS and stained by adding100 μl of staining solution (1 ml solution contains 973 μl of PBS, 8 μl of 0.5M potassium ferrocyanide, 8 μl of 0.5M potassium ferricyanide, 1 μl of 2M MgCl2 and 10 μl of 40 mg/ml X-gal) and incubating overnight at 37°C. Staining was stopped by the removal of the staining solution and washing twice with PBS. The number of blue stained, infected cells was counted with a Nikon TS100-F inverted microscope and the infectivity was expressed as the number of infected cells per 100 pg p24 antigen.
The levels of the p24 ELISA observed and the infectivity counts were represented as graphs plotted using Excel. Significance of differences between viral strains were determined through statistical analysis by use of the Statistical Package for Social Sciences (SPSS) software version 20. In view of the limited numbers, comparison of virion productivities and viral infectivities were done by the nonparametric Kuskal-wallis one way ANOVA test.
Ethical approval was obtained from the Institutional Review Board of the Noguchi Memorial Institute for Medical Research of the College of Health Sciences, University of Ghana. Voluntary informed consent was obtained from the study participants.
Virion productivity and infectivity of prevalent HIV strains in Ghana were examined. Virions were prepared from cells transfected with proviral DNA clones, and their infectivity was determined by MAGI assay, a single-cycle infection assay. In Table 1 the viral strains tested and the initial viral isolation done are shown. Initial viral isolation was successful for all samples on MAGIC5A cells but not all were successful on peripheral blood lymphocytes. To allow for comparison, 8 viral isolates comprising 3 subtype A, 2 subtype G and 3 recombinant subtype CRF02_AG that were isolated on both PBL and MAGIC5A cells were selected for virion productivity and infectivity assays.
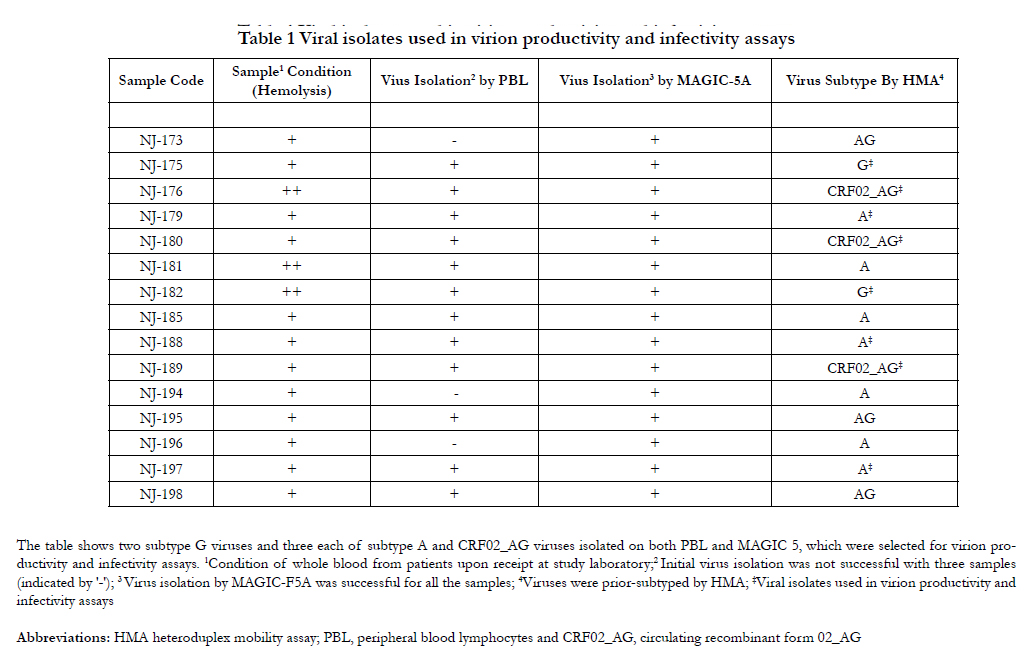

Table 1 Viral isolates used in virion productivity and infectivity assays
HIV-1 p24 ELISA, an enzyme immunoassay was performed to determine the concentration of HIV-1 p24 produced in the cell culture samples. The HIV -1 p24 core antigen is a component of the various viral proteins that combine with the two RNA strands to form the virion core. Since the amino acid sequence of HIV-1 p24 is well conserved among a number of HIV-1 isolates, p24 assay detects various isolates with comparable sensitivity. The assay results of the isolates studied were within the linear range of the assay and the concentrations obtained are shown in Figure 1. Measurement of p24 antigens show viral particles were produced in all transfected 293T cells. The levels of the p24 production observed indicate that virion productivity was almost comparable; that is, there was no significant difference (p =0.405) in virion productivity among the viral subtypes tested.
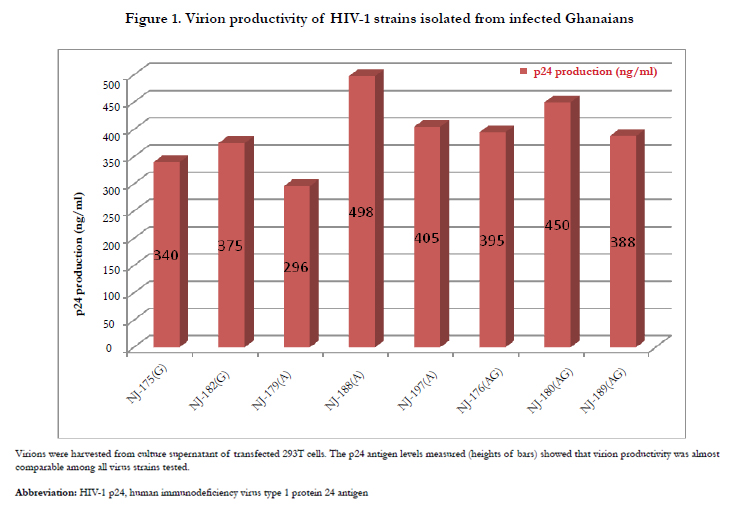
Figure 1. Virion productivity of HIV-1 strains isolated from infected Ghanaians.
The MAGIC5A cells used in the infectivity assay are cells engineered to express CD4 receptor and co-receptors CXCR4 and CCR5. The cells are therefore infectable by the HIV-1 isolates studied, whether the infecting viruses are of X4, R5 or dual tropism. Also, MAGIC5A cell line used had been engineered to augment CD4 expression since primary HIV-1 has been reported to depend on CD4 density more than laboratory strains of HIV-1 such as HIV-1NL432 [12, 16, 17]. The genetically engineered MAGIC5A cell contains a virus-inducible reporter gene cassette comprising the viral long terminal repeat (LTR), placed upstream of the gene encoding E. coli β-galactosidase (lacZ). Expression of the lacZ gene is therefore dependent upon the transcriptional activity of the HIV LTR. The expression activity is known to be weak, that is, β-galactosidase produced under normal circumstances is too little to detect. However, detectable β-galactosidase production is greatly enhanced if the cell becomes infected by HIV-1 and the virus produces its transcriptional activator protein, Tat, which in turn upregulates the activity of the viral LTR [18].The expression of the enzyme β-galactosidase is detected in the MAGI assay procedure by use of the chromogenic substrate, X-gal contained in the staining solution. The level of expression is therefore in proportion to the infecting virions.
Given the above indicator cell properties, HIV virion infectivity was determinable by the MAGI assay. Figure2. shows the microscopic. results of the assay. Majority of blue cells were singles and each was counted as resulting from one infectious viral particle. However, occasionally, blue cells appeared in clusters presumably due to cell division and therefore a cluster of blue cells was counted as resulting from one infectious particle. A more detailed graphical representation of the levels of infectivity of the HIV-1 isolates studied are shown in Figure3. Infectivity has been expressed as the number of infected cells per 100 pg p24 antigen. In view of the limited number of strains, the infectivity levels were compared statistically using the non-parametric test of Kruskal-Walis One way ANOVA. The analysis showed significant difference (p=0.044) between levels of infectivity of viral subtypes with subtype CRF02_AG showing significantly higher infectivity.
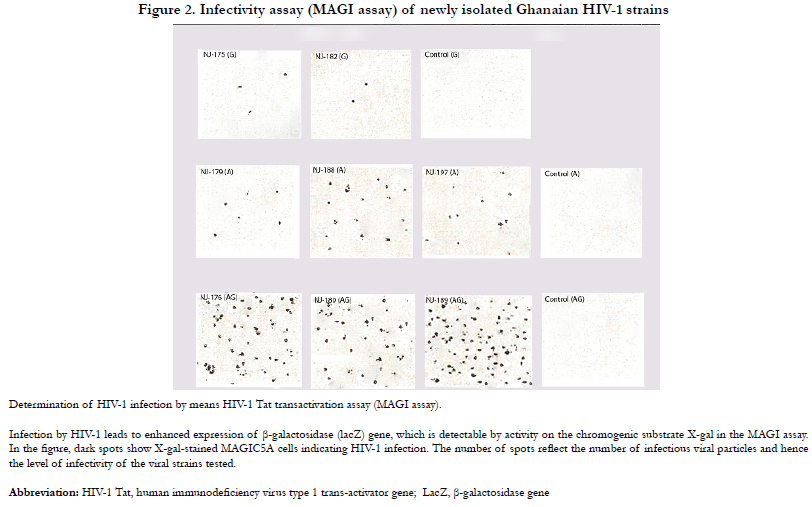
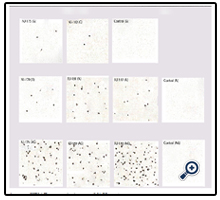
Figure 2. Infectivity assay (MAGI assay) of newly isolated Ghanaian HIV-1 strains.
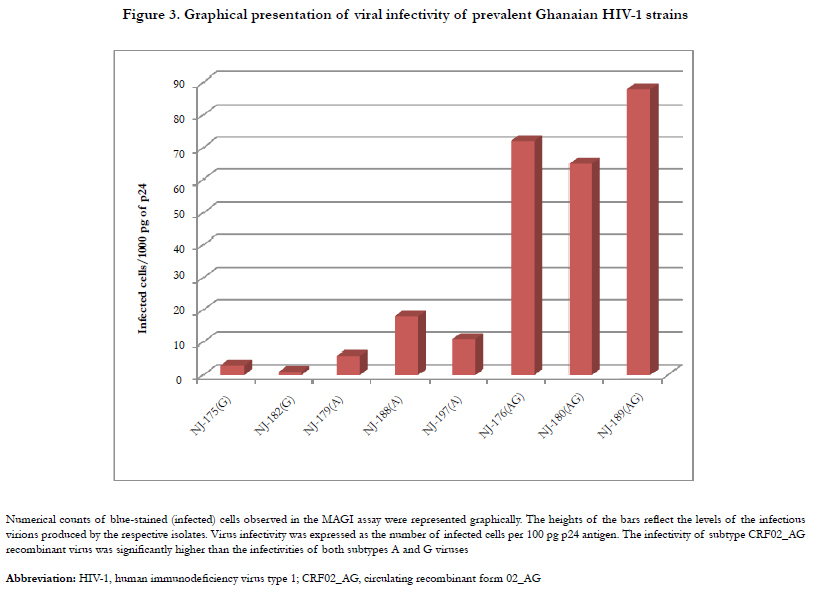
Figure 3. Graphical presentation of viral infectivity of prevalent Ghanaian HIV-1 strains.
Discussion
This study examined virion productivity and viral infectivity of prevalent HIV-1 strains in Ghana and found a recombinant strain CRF02_AG to have higher infectivity than the parental subtypes A and G. Virion productivity was assessed through cell transfection. The results of the p24 ELISA indicate transfection success and virus production. In this assay, the optical density values are proportional to the amount of HIV-1 p24 antigen in the viral specimen. However, even though virus productivity was almost comparable among all strains tested, this did not reflect in the infectivity test outcome.
Sixteen hours post-transfection culture medium was changed; and viruses were harvested after 48 hrs post-transfection. This is required for the transfection procedure employed in order to maintain healthy cell growth and enhance viral infectivity [14, 19]. Infectivity in this study refers to the number of virions that could establish productive infection in host indicator cell line. Infection of the cells for two hours at 37°C and further incubation for 48 hours was to allow production of β-galactosidase in infected cells that could be visualized by X-Gal staining in which infected MAGIC5A cells appear in blue color [20]. Counting of stained cells under the microscope therefore gave the number of cells that had been productively infected by varions and which produced Tat protein upon transcriptional activation, necessary for the activated expression of β-galactosidase. Appearance of more blue stained cells of the CRF02_AG test suggests this strain is more infectious in vitro than the other strains tested.
The observation of CRF02_AG recombinant viruses displaying comparatively higher infectivity than those of both subtypes A and G combined might reflect what occurs in vivo in the Ghanaian situation, which may explain the continued dominance of the CRF02_AG recombinant for well over a decade now [21-23]. The apparently lower infectivity of the parental subtypes A and G may either reflect an intrinsically low infectivity of the virions themselves or perhaps due to an unknown viral-cell interactions that restrict HIV-1 infection of those strains. However, it could be assumed that a large number of the viral particles produced were defective, hence virion productivity did not correspond with productive viral infection in the parental strains. Attempt was made to compare a consistent number of circulating strains but 3 subtype A, 2 subtype G and 3 CRF02_AG recombinant isolates were obtained for comparison. Nevertheless, the difference in infectivity between the CRF02_AG and the other pure subtypes tested (p=0.044) may suggest the influence of recombination; that is, recombination might give advantage of higher infectivity.
Recombination might arise due to natural evolution of strains; it might not be the predominant event that explains the difference in infectivity. Other factors may also play a role in determining the outcome of productive viral infection. For example, with its limited genome size, HIV is known to interact with both supportive and inhibitory host factors in undergoing its life cycle [24-26]. Many viral evolutionary mechanisms therefore evolve counteracting strategies to neutralize the activities of host cellular restriction factors [27, 28]. Thus even though virion infectivity could be enhanced by reduction of defective virions in a virus pool through variation of culture conditions such as medium change, the unknown difference in virus-cell interactions may be a factor that restricts HIV-1 virion infectivity [29].This study did not undertake to determine the role of factors that might potentially influence viral infectivity.
To date, inhibition of viral replication remains an important theme in the ongoing fight against HIV. In the absence of anti- HIV vaccine, current treatment of HIV/AIDS is by use of antiretroviral therapy (ART) drugs, which either inhibit viral entry or target viral proteins reverse transcriptase, protease and integrase [30, 31]. It is noteworthy that in both treated and untreated HIV infection accurate and sensitive assessment of viral load remains essential in initiating, monitoring response and in considering switching ART regimens [32].This study therefore highlights the potential relevance of in vitro measurements of viral infectivity in the management of in vivo behavior of HIV-1 infection.
This study is discussed in the light of some limitations. The number of strains available for comparison were limited, in which case comparison of recombinant viruses would require doing so only to their direct parents. Importantly, due to the natural divergence of strains, comparing a limited number of strains might result in comparing strains that evolved from each other many years or decades before. Furthermore, since expression of β-galactosidase depends on the upregulation of the viral LTR activity by the Tat protein, sequence variation in the LTR region may influence the Tat activation readout and thereby impact the MAGI assay. Nevertheless, the findings of this in vitro infectivity study shed some light on the epidemiological significance of the strains tested in the in vivo situation.
Conclusion
This study examined production of virions from cells transfected with proviral DNA clones of isolated HIV-1 strains and determined their infectivity by the MAGI assay. No significant difference was detected in the levels of virion production of the viruses in transfected cells but viral infection assay showed the infectivity of a recombinant strain, CRF02_AG, higher than both parental subtypes A and G. Thus virulence of the infecting HIV- 1 subtype and presumably the kinetics of viral-host interactions might represent a considerable factor in defining the clinical course of HIV-1 infection. Understanding determinants of HIV-1 infectivity and the factors that influence viral replication could prove useful in the search for strategies for new therapeutic interventions.
Acknowledgements
We would like to thank the Japan International Cooperation Agency (JICA) for their support. Most part of the work was conducted through support from JICA with funds for travelling and stay in Japan as part of its efforts to provide capacity building in developing countries. The National Institute for Infectious Diseases (NIID), Tokyo, Japan supported the work with reagents and lab space while the Virology Department of the Noguchi Memorial Institute for Medical Research supported with initial sample processing.
References
- Leland DS, Ginocchio CC (2007) Role of cell culture for virus detection in the age of technology. Clin Microbiol Rev 20(1): 49–78.
- Flint JS, Enquist LW, Shalka AM (2015) Virological Methods. In: Principles of Virology. http://www2.oakland.edu/biology/chaudhry/pics/Virological-Methods.pdf. Accessed 5th .
- Olivo PD (1996) Transgenic cell lines for detection of animal viruses. Clin Microbiol Rev 9(3): 321–334.
- Freed EO, Martin MA (2013) Human Immunodeficiency Viruses: Replication. In: Fields Virology: Lippincott. Edited by Knipe DM, Howley PM. Williams and Wilkins; p1502–1560.
- Clapham P, McKnight A (2002) Cell surface receptors, virus entry and tropism of primate lentiviruses. J Gen Virol 83(8): 1809–1820.
- Anderson J, Akkina R (2005) TRIM5alpharh expression restricts HIV-1 infection in lentiviral vector-transduced CD34+-cell-derived macrophages. Mol Ther 12(4): 687–696.
- Huthoff H, Towers GJ (2008) Restriction of retroviral replication by APOBEC3G/F and TRIM5α. Trends Microbiol 16(12): 612–619.
- Mansky LM, Temin HM (1995) Lower in vivo mutation rate of human immunodeficiencyvirus type 1 than that predicted from the fidelity of purified reverse transcriptase. J Virol 69(8): 5087–5094.
- Korber B, Muldoon M, Theiler J, Gao F, Gupta R, et al. (2000) Timing the ancestor of the HIV-1 pandemic strains. Science 288(5472): 1789–1796.
- Simon V, Ho DD, Abdool KQ (2006) HIV/AIDS epidemiology, pathogenesis, prevention, and treatment. Lancet 368(9534): 489–504.
- Bushman FD, Nabel GJ, Swanstrom R (2012) HIV - from biology to prevention and treatment. Cold Spring Harbor,Cold Spring Harbor Laboratory Press,New York. p. 572.
- Mochizuki N, Otsuka N, Matsuo K, Shiino T, et al. (1999) An infectious DNA clone of HIV type 1 subtype C. AIDS Res Hum Retrovir 15(4):1321–1324.
- Hachiya A, Aizawa-Matsuoka S, Tanaka M, Takahashi Y, Ida S, et al. (2001) Rapid and simple phenotypic assay for drug susceptibility of human immunodeficiency virus type 1 using CCR5-expressing HeLa/CD4+ cell clone 1–10 (MAGIC-5). Antimicrob. Agents Chemother 45(2): 495–501.
- Kingston RE, Chen CA,Rose JK (2003) Calcium phosphate transfection. Current protocols in molecular biology, 9-1.
- Wigler M, Pellecer A, Silverstein S, Axel R, Urlaub G, et al. (1979) DNAmediated transfer of the adenine phosphoribosyltransferase locus into mammalian cells. Proc Nat Acad Sc USA 76(3): 1372–1376.
- Kozak EJ, Platt SL, Kabat D (2000) Critical role of enhanced CD4 affinity in laboratory adaptation of human immunodeficiency virus type 1. AIDS Res Hum Retrovir 16(9): 871–882.
- Kozak SL, Platt EJ, Madani N, Ferro Jr FE, Peden K, et al. (1997) CD4, CXCR-4, and CCR-5 dependencies for infections by primary patient and laboratory-adapted isolates of human immunodeficiency virus type 1. J Virol 71(2): 873–882.
- Brass AL, Dykxhoorn DM, Benita Y, Yan N, Engelman A, et al. (2008) Identification of host proteins required for HIV infection through a functional genomic screen. Science 319(5865): 921–926.
- Berkhout B, Haasnoot J (2006) The interplay between virus infection and the cellular RNA interference machinery. FEBS Lett 580(12): 2896–2902.
- Kimpton J, Emerman M (1992) Detection of replication-competent and pseudotype human immunodeficiency virus with a sensitive cell line on the basis of activation of an integrated-galactosidase gene. J Virol. 66(4):2232–2239.
- Fischetti L, Opare-Sem O, Candotti D, Sarkodie F, Lee H, et al. (2004) Molecular epidemiology of HIV in Ghana: Dominance of CRF02_AG. J Med Virol. 73(2): 158–166.
- Delgado E, Ampofo WK, Sierra M, Torpey K, Pérez-Alvarez L, et al. (2008) High prevalence of unique recombinant forms of HIV-1 in Ghana: molecular epidemiology from an antiretroviral resistance study. J Acquir Immun Defic Syndr 48(5): 599–606.
- Nii-Trebi NI, Ibe S, Barnor JS, Ishikawa K, Brandful JAM, et al. (2013) HIV-1 Drug-Resistance Surveillance among Treatment-Experienced and -Naı¨ve Patients after the Implementation of Antiretroviral Therapy in Ghana.PLoS One 8(8): e71972.
- Adamson CS, Freed EO (2010) Novel approaches to inhibiting HIV-1 replication. Antiviral Res 85(1): 119–141.
- Bushman FD, Malani N, Fernandes J, D’Orso I, Cagney G, et al. (2009). Host cell factors in HIV replication: meta-analysis of genome-wide studies. PLoS Pathog 5(5): e1000437.
- Liu L, Oliveira NM, Cheney KM, Pade C, Dreja H, et al. (2011) A whole genome screen for HIV restriction factors. Retrovirology 8: 94.
- Friedrich BM, Dziuba N, Li G, Endsley MA, Murray JL, et al. (2011) Host factors mediating HIV-1 replication (Review). Virus Res 161(12): 101–114.
- Freed EO (2004) HIV-1 and the host cell: an intimate association. Trends Microbiol. 12(4): 170–177.
- Zheng Y-H, Jeang K-T, Tokunaga K (2012) Host restriction factors in retroviral infection: promises in virus-host interaction. Retrovirology 9: 112.
- De Béthune MP (2010) Non-nucleoside reverse transcriptase inhibitors (NNRTIs), their discovery, development, and use in the treatment of HIV-1infection: A review of the last 20 years (1989–2009). Antiviral Res 85(1):75–90.
- Tilton JC, Doms RW (2010) Entry inhibitors in the treatment of HIV-1 infection. Antiviral Res 85(1): 91–100.
- Walker BD, Yu XG (2013) Unraveling the mechanisms of durable control of HIV-1. Nat Rev Immunol 13(7): 487–498.





