How Zika Virus Crosses The Placental Barrier?
Shafique S*
Anatomical Sciences Program, Department of Biomedical and Molecular Sciences, Queen’s University, Kingston, Ontario, Canada.
*Corresponding Author
Sidra Shafique
Anatomical Sciences Program, Department of Biomedical and Molecular Sciences,
Queen’s University, Kingston, Ontario, Canada.
Tel: 1-613-533-2727
Fax:1-613-533-2022
E-mail: s.shafique@queensu.ca
Received: July 20, 2016; Accepted: September 23, 2016; Published: September 26, 2016
Citation: Shafique S (2016) How Zika Virus Crosses The Placental Barrier?. Int J Vaccines Res. 3(4), 31-34. doi: dx.doi.org/10.19070/2572-7427-160006
Copyright: Shafique S© 2016. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
The epidemic of Zika Virus (ZV) drastically affected the health of more than one million people in Brazil resulting in twenty-fold increase of microcephaly (head circumference of newborn baby being less than or equal to thirty-two cms) in infected pregnant women. ZV crosses the placental barrier between maternal and fetal compartments. ZV has been found in amniotic fluid and fetal brain tissue of the human pregnancies complicated with ZV infection and microcephaly supporting cause and effect relationship between ZV and microcephaly. New knowledge about how ZV enters into the cells is mandatory for preventive and therapeutic research. ZV is a Flavivirus, therefore the Flavivirus receptors are candidate receptors for ZV too. ZV has been associated with high level of TYRO3 and AXL component of TAM, a Tyrosine Kinase receptor family, expression in the infected skin cells and neuronal stem cells. It predicts that AXL receptors may facilitate ZV entry into the trophoblasts and endothelial cells lining the fetal vessels present in the placenta. This hypothesis stems from the reported studies on ZV and AXL receptor expression, both in human tissues and animal models. If our hypothesis proves to be valid, it would be a great step towards the drug and therapeutic development to prevent ZV crossing the placental barrier and resulting in complications.
2.Introduction
3.Why Zika Virus is so important?
4.Zika Virus and Placental Barrier
5.AXL as Zika Virus Entry Receptors
6.Conclusion
7.Future Directions
8.Acknowledgements and Conflict of Interest
9.References
Keywords
Zika Virus; Placental Barrier; Microcephaly; AXL Receptors; Flavivirus Receptors.
Introduction
The epidemic of Zika Virus (ZV) started in April 2015 in Brazil and since then more than one million individuals have been affected by it [1]. ZV was named after the forest in Uganda where it was first found, in 1947 [2]. Phylogenetically, it is related to Spondweni virus clade, an arthropode-borne virus transmitted by mosquitoes, the family being Flaviviridae and the genus Flavivirus [3,21]. Two lineages of this virus include the African lineage, identical to the strain spreading in South America and, the Asian lineage identified in the 2013-2014 outbreak of French Polynesia [4]. A mosquito, Anopheles Aegypti, is the biological vector responsible for its spread [1]. Sexual transmission of ZV has been documented in WHO report of April 2016 in United States, Italy, France, Chile, Argentina and New Zealand [19, 20]. Like other Flaviviruses, ZV is an enveloped, icosahedral virus, having single-stranded, positive-sense RNA genome that can be translated into viral proteins [3]. 3.7°A resolution cryoelectron microscopy reveals its resemblance to other Flaviviruses in architecture but comparison of the structure of its envelope (E) protein indicates that some parts of this protein are similar to West Nile and Japanese encephalitis viruses, while other components are more like Dengue virus [22].
ZV is more stable at higher temperatures, 40°C and above, than Dengue virus strains, DENV2 and DENV4, and this characteristic seems to contribute its high infectivity at human body temperature (37°C) and high climatic temperatures of Brazil [22]. Unlike other Flaviviruses, ZV can cross the placenta through unknown mechanisms. ZV has been found almost all over the globe from Africa to Southeast Asia, but remained out of focus due to its sporadic epidemics, for example outbreak in Yap island, Micronesia affected only fourteen people in 2007 [26]. The epidemic of Brazil involving South America is so far its largest and rapid spread.
Why Zika Virus is so important?
ZV causes fever and rash, neurological complications including Guillain-Barre (GB) syndrome during French Polynesia outbreak where twenty fold rise in incidence of GB syndrome is reported [10, 27]. In non-pregnant women, it usually causes mild flu-like symptoms while is a risk factor for the developing conceptus in infected pregnant women.
Zika virus has been associated with ocular complications and microcephaly (head circumference of newborn baby being less than or equal to thirty two cms) in infected pregnant women [6, 8]. ZV crosses the placental barrier at maternal-fetal interface, proliferates in the amniotic fluid and damages the developing brain tissue [7, 9, 10]. In Brazil, ZV has been associated with an increased incidence of microcephaly to such an extent that by January 2016, the Brazilian Ministry of Health had reported 3174 microcephalic newborns, corresponding to a 20-fold increase over the expected annual reported incidence of microcephaly [5, 6]. Zika is a teratogenic virus. It has the potential to cause anomalies in the developing fetus and its brain resulting in permanent damage to neuronal tissue and irreversible ocular destruction when maternal infection occurs in first or early second trimester maternal infection [8]. ZV RNA has been identified in the amniotic fluid of infected pregnant women with microcephalic fetuses and in the brain tissue of microcephalic fetuses as well, indicating a cause and effect relationship between Zika virus and microcephaly [7]. Other reported complications include placental insufficiency with fetal growth retardation, fetal death or abortion being the extreme complications [5]. Therefore, it is important to investigate the research question ‘How does the ZV crosses the placenta and infects the fetus?’ Answers will prevent further destruction of humans in the current generation and will prepare us to deal with future epidemics. Animal models may play important roles in finding the answer to the question. It is already known that the Asian strain SZ01 infects mouse brain tissue in vivo and affects mouse fetal brain development subsequent to crossing through the placenta [9,10].
Zika Virus and Placental Barrier
The placenta is the organ that supplies nutrients to and removes wastes from the developing fetus. Maternal and fetal circulations are separated only by the placental barrier [11]. This barrier is formed by syncytiotrophoblasts and endothelial cells lining the fetal vessels in species having hemochorial placentas such as humans and mice [11].
Mice are used extensively to model human placental diseases due to these similarities in placental structure and function [12]. The hemochorial and discoid mouse placenta consists of the maternal decidua and conceptus-derived components, the junctional and labyrinth zones of trophoblast, an epithelium-derived cell lineage. Conceptus-derived trophoblast giant cells, glycogen trophoblasts and spongiotrophoblasts are different types of trophoblast cells present in all three zones of the mouse placenta. In the labyrinth zone, the fetal capillaries are lined by the endothelium that is covered by a layer of mononuclear trophoblasts and syncytiotrophoblasts [11]. ZV must infects the trophoblast cells before it can enter the fetal compartment [12]. In the ZV susceptible IFNAR null mouse, placental cellular damage follows infection, the labyrinth zone becomes markedly thinned as seen by reduction in immunostaining with the pan-trophoblast/ epithelial cell marker, pan-cytokeratin [10]. Transmission electron microscopic images of these mouse placentas revealed 50nm virions in the endoplasmic reticulum of both trophoblast cells and in the fetal vascular endothelial cells of the placental labyrinth [10]. In ZV infected IFNAR null mouse placenta, vimentin a stain for, fetal blood vessels, is decreased, reflecting fetal endothelial cell damage by the virus [10]. These findings support the hypothesis that ZV crosses the placental barrier by infecting and destroying the trophoblast cells of this barrier and then infecting the fetal endothelium. Damage to this cell type or replication within it would permit ZV to enter the systemic fetal blood stream via the umbilical cord circulation. This would explain why the amnion is not protective against the virus. It is possible that the hematogenous spread of ZV in the fetus leads to cutaneous fetal lesions that shed virus into the amniotic cavity. Systemic circulation of ZV would permit the neurotrophic properties of the virus to be expressed and the fetal brain to become infected.
The use of the mouse model for the study of mechanisms of ZV placental infection is valid because cultured human trophoblast cell lines are reported to be infected and damaged in the same manner [12]. ZV RNA is found in the human placentas of human fetuses with anomalies [7]. In humans maternal blood starts circulating in intervillous space of the developing placenta around eight to ten weeks and symptoms of ZV infection have been reported as early as eight weeks of gestation in women delivering microcephalic babies. Thus, maternal placental circulation disseminates ZV and contributes to its teratogenesis [28]. It is also possible that ZV may become part of immune complexes and be actively transported by receptors across the placenta. The mouse placenta experiments favors the ZV crossing of the placenta as a sequelae of infecting placental barrier cell line rather than using diffusion or cargo processes [28].
AXL as Zika Virus Entry Receptors
Molecular mechanisms must be identified for ZV entry into the cells. Flaviviruses infect the cells by interacting a defined set of receptors that facilitate endocytosis of virus particles [15].Flavivirus receptors are anticipated to facilitate ZV entry into cells, as these are induced and over-expressed in infected mouse fetal brains [9]. Out of these, the phosphatidylserine receptors including T-cell immunoglobulin and mucin domain (TIM) and TYRO3, AXL and MER (TAM) are actively expressed in ZV infected cells [13] [Figure 1]. Zika and dengueviruses both infect human skin epithelial cells via AXL receptors [13]. Phosphatidylserine (PtdSer) is a cell membrane phospholipid that normally faces towards the cytosol of a cell. When there is cell damage, it is exposed to cells’ exterior and acts as a signal for specific receptors to stimulate endocytosis. Same PtdSer molecule is expressed on ZV surface [24]. Although both TIM and TAM receptors have been active in skin fibroblasts and immune cells to help the ZV enter in the human skin cells after an infectious mosquito bite, but TAM receptors are more favoured as ZV receptors because the AXL component of this complex is over-expressed in ZV-infected human and mouse fetal brains after ZV infection [9, 13, 16]. The TAM receptor complex has been grouped in the tyrosine kinase family since 1991 [24]. Gas6 and Protein S are their two ligands that are present on the receptor complex and recognizes the viral PtdSer [15]. Viral PtdSer attachment to these ligands stimulates the process of endocytosis followed by a signaling cascade [15]. The AXL component of the TAM receptor is a Tyrosine kinase, coded by AXL gene and is an active component of TAM complex [15]. This receptor is present in developing neural tissue and radial glia; expression of AXL being conserved in animal models of cortical development [16]. Experiments with mouse model have shown that the AXL receptor gene is over-expressed along with microcephaly genes after ZV infection in E13.5 to E16.5 fetal brains. This is accompanied by a three-hundred-fold increase of viral RNA copies in the neural progenitor cells resulting in microcephaly [9].
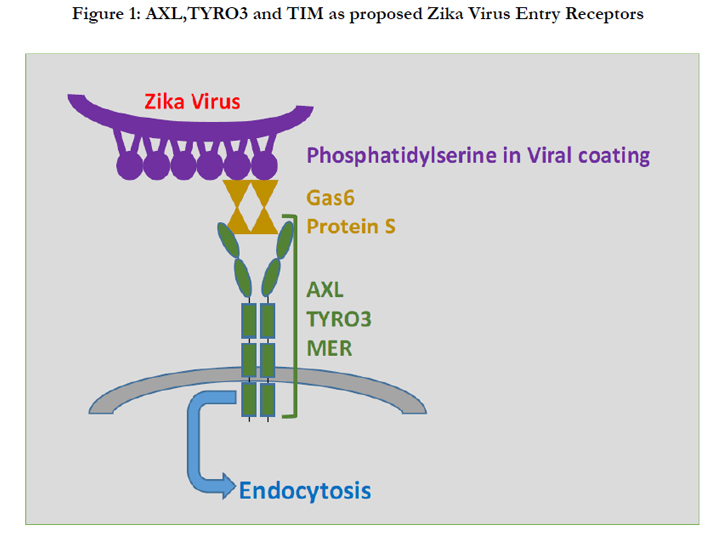
Figure 1: AXL,TYRO3 and TIM as proposed Zika Virus Entry Receptors.
Conclusion
Zika Virus belongs to the Flavivirus family of viruses while AXL receptors are Flavivirus family receptors that are induced by ZV infection of skin fibroblasts and the neuronal progenitor cells [9]. In infected mouse brains, over expression of AXL protein accompanies induction of microcephaly genes and the gross anatomical observation of microcephaly. AXL receptors are induced in the skin cells where the mosquito bites inoculate ZV [13]. Thus my hypothesis is that AXL is expressed by trophoblast cells and contributes to their infection by ZV. Existing data suggest that ZV infects mouse and human trophoblasts disrupting the outermost layer of the maternal-fetal barrier. ZV also infects and damages the fetal vascular endothelial cells of the placenta by subsequent direct infection [10, 11] Evidence is yet to be presented in the literature on whether ZV receptors are expressed by or induced in trophoblasts. If this (my) hypothesis is correct, it would identify a target for the development of a therapeutic strategy for prevention of ZV transmission to developing fetuses in pregnant women.
Study approaches for ZV receptors may include ifnar-/- mouse model or the TEM receptor knock out mouse models. [10, 24]. Human placenta cell lines and human placenta models are increasingly being used to study human microbial pathogenesis [29]. Human placental cytotrophoblasts and fetal Hofbauer cells are not only susceptible to ZV infection in vitro but there is sufficient evidence for ability of ZV to replicates in these cells causing minimal cell death [29]. The progression of ZV pathogenesis in placenta may largely depends on this factor, since minimal cell death releases minimal cellular debris thus low levels of these inflammatory signals help the virus to hide and escape causing devastating tissue damage [29]. Recently, a new threedimensional villus formation human placenta model was created that may help to understand human fetal-maternal interface in more detail for vertical transmission of infectious agents such as ZV [30].
Future Directions
New knowledge on the mechanisms of ZV infection is urgently required. Flaviviruses are typically unable to cross the placenta but ZV is an exception using unknown mechanisms. Evidence based data about the ZV receptors will help to develop the therapeutic or preventive strategies regarding its teratogenic effects on human fetuses. The available literature supports the hypothesis that ZV infects the fetus by crossing the placental barrier and infecting fetal cells rather than through any remote inflammatory mechanism. The human placental barrier is complex and has many mechanisms to protect the fetus until birth. Discovering the mechanism(s) used by ZV to evade evolutionary conceptus protection will provide a break through towards the development of therapeutic approaches to prevent ZC’s teratogenic effects in the future. Epidemiologists, pathologists, virologists and biologists with expertise in placenta research are working towards this goal and hope to soon achieve better outcomes for ZV exposed human pregnancies.
Acknowledgements and Conflict of Interest
The author is acknowledged to Dr. B. Anne Croy, DVM, Ph.D. Tier 1 Canada Research Chair in Reproduction, Development and Sexual Function. Professor, Department of Biomedical and Molecular Sciences, Queen’s University, Kingston. ON. For reviewing the content of this manuscript. There is no conflict of interest whatsoever related to the submitted material for publication.
References
- Hennessey M, Fischer M, Staples JE (2016) Zika virus spreads to new areas—region of the Americas, May 2015–January 2016. American Journal of Transplantation. 16(3): 1031-1034.
- Malone RW, Homan J, Callahan M V, Glasspool-Malone J, Damodaran L,et al., (2016) Zika virus: medical countermeasure development challenges. PLoS Negl Trop Dis.10(3): e0004530.
- Faye O, Freire CC, Iamarino A, de Oliveira JVC, Diallo M, et al., (2014) Molecular evolution of Zika virus during its emergence in the 20 th century. PLoS Negl Trop Dis. 8(1): e2636.
- Enfissi A, Codrington J, Roosblad J, Kazanji M, Rousset D (2016) Zika virus genome from the Americas. The Lancet. 387(10015): 227-228.
- Ventura CV, Maia M, Ventura B V, Linden VV D, Araújo EB, et al., (2016) Ophthalmological findings in infants with microcephaly and presumable intra-uterus Zika virus infection. Arquivos brasileiros de oftalmologia. 79(1):1-3.
- De Paula Freitas B, de Oliveira Dias JR, Prazeres J, Sacramento GA, Ko AI, et al., (2016) Ocular findings in infants with microcephaly associated with presumed Zika virus congenital infection in Salvador, Brazil. JAMA ophthalmology. 134(5): 529-535.
- Calvet G, Aguiar RS, Melo AS, Sampaio SA, de Filippis I, et al., (2016) Detection and sequencing of Zika virus from amniotic fluid of fetuses with microcephaly in Brazil: a case study. The Lancet Infectious diseases.16(6):653-660.
- Jampol LM, Goldstein DA (2016) Zika virus infection and the eye. JAMA Ophthalmology. 134(5): 535-536.
- Li C, Xu D, Ye Q, Hong S, Jiang Y, et al., (2016) Zika virus disrupts neural progenitor development and leads to microcephaly in mice. Cell stem cell.
- Miner JJ, Cao B, Govero J, Smith AM, Fernandez E, et al., (2016) Zika virus infection during pregnancy in mice causes placental damage and fetal demise. Cell. 165(5): 1081-1091.
- Simmons DG, Cross JC (2005) Determinants of trophoblast lineage and cell subtype specification in the mouse placenta. Dev Biol. 284(1): 12-24.
- Bayer A, Lennemann NJ, Ouyang Y, Bramley JC, Morosky S, et al., (2016) Type III interferons produced by human placental trophoblasts confer protection against Zika virus infection. Cell Host Microbe. 19(5): 705-712.
- Hamel R, Dejarnac O, Wichit S, Ekchariyawat P, Neyret A, et al., (2015) Biology of Zika virus infection in human skin cells. J virol. 89(17): 8880- 8896.
- Adibi JJ, Marques ET, Cartus A, Beigi RH (2016) Teratogenic effects of the Zika virus and the role of the placenta. Lancet. 387(10027): 1587-1590.
- Perera-Lecoin M, Meertens L, Carnec X, Amara A (2013) Flavivirus entryreceptors: an update. Viruses. 6(1): 69-88.
- Nowakowski TJ, Pollen AA, Di Lullo E, Sandoval-Espinosa C, Bershteyn M,vet al., (2016) Expression analysis highlights AXL as a candidate Zika virus entry receptor in neural stem cells. Cell stem cell. 18(5): 591-596.
- Hwang S Y, Hertzog PJ, Holland KA, Sumarsono SH, Tymms MJ, et al., (1995) A null mutation in the gene encoding a type I interferon receptor component eliminates antiproliferative and antiviral responses to interferons alpha and beta and alters macrophage responses. Proc Natl Acad Sci. 92(24): 11284-11288.
- Baronti C, Piorkowski G, Charrel RN, Boubis L, Leparc-Goffart I, (2014) Complete coding sequence of Zika virus from a French Polynesia outbreak in 2013. Genome Announc. 2(3): e00500-14.
- Oster Alexandra M, Russell Kate, Stryker Jo Ellen, Friedman A, Kachur RE,et al., (1 April 2016) Update:Interim Guidance for Prevention of Sexual Transmission of Zika Virus. MMWR. Morb Mortal Wkly Rep. 65 (12):323–25.
- "Zika Virus Microcephaly And Guillain-Barré Syndrome Situation Report" .World Health Organization. 7 April 2016.
- Chambers Thomas J, Monath Thomas P (2003)The Flaviviruses: Detection, Diagnosis and Vaccine Development: Detection, Diagnosis and Vaccine Development, Academic Press, 376.
- Kostyuchenko V A, Lim EX, Zhang S, Fibriansah G, Ng TS, et al., (2016) Structure of the thermally stable Zika virus. Nature.
- Verhoven B, Schlegel R, Williamson P (1995) Mechanisms of phosphatidylserine exposure, a phagocyte recognition signal, on apoptotic T lymphocytes. J Exp Med: 182(5): 1597-1601.
- Lemke G (2013) Biology of the TAM receptors. Cold Spring Harb perspect- Biol. 5(11): a009076.
- Prasad D, Rothlin CV, Burrola P, Burstyn-Cohen T, Lu Q, et al., (2006) TAM receptor function in the retinal pigment epithelium. Molecular and Cellular Neuroscience. 33(1): 96-108.
- Duffy MR, Chen TH, Hancock WT, Powers AM, Kool JL, et al., (2009) Zika virus outbreak on Yap Island, federated states of Micronesia. New Eng and Journal of Medicine. 360(24): 2536-2543.
- Oehler E, Watrin L, Larre P, Leparc-Goffart I, Lastere S, et al., (2014) Zika virus infection complicated by Guillain-Barre syndrome--case report, French olynesia, December 2013. Euro Surveill. 19(9): 20720.
- Foy BD, Kobylinski KC, Chilson Foy JL, et al., (2011) Probable non-vectorborne transmission of Zika virus, Colorado, USA. Emerg Infect Dis. 17:880–82.
- Quicke KM, Bowen JR, Johnson EL, McDonald CE, Ma H, et al., (2016) Zika Virus Infects Human Placental Macrophages. Cell Host Microbe. 20(1): 83-90.
- “New Placenta Model Could Reveal How Birth Defect-Causing Infectious Agents Cross From Mother to Baby”, Pittsburgh, March 4, 2016.





