Ragusana Donkey Milk as a Source of Lactic Acid Bacteria and Yeast Strains of Dairy Technological Interest
Randazzo CL1, Restuccia C1, Mancini A2,3, Muccilli S1, Gatti M2,4, Caggia C1*
1 Department of Agriculture, Food and Environment (Di3A), University of Catania, Catania, Italy.
2 Department of Food Science, University of Parma, Parma, Italy.
3 Nutrition and Nutrigenomics group, Department of Food Quality and Nutrition, Research and Innovation Centre, Fondazione Edmund Mach di San
Michele all’Adige, Trento, Italy.
4 Multidisciplinary Interdepartmental Dairy Center (MILC), University of Parma, Italy.
*Corresponding Author
Cinzia Caggia,
Department of Agriculture, Food and Environment (Di3A),
University of Catania, via S. Sofia 98, 95123 Catania, Italy.
Tel: +(39)0957580218
E-mail: ccaggia@unict.it
Received: April 21, 2016; Accepted: May 17, 2016; Published: May 24, 2016
Citation: Caggia C, et al., (2016) Ragusana Donkey Milk as a Source of Lactic Acid Bacteria and Yeast Strains of Dairy Technological Interest. Int J Dairy Process Res. 3(2), 38-46.
Copyright: Caggia C© 2016. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
Donkey milk is considered an interesting substitute to cow’s milk thanks to its nutritional properties. The chemical composition of donkey milk has been extensively considered essential to prevent the growth of undesirable microorganisms, and few data are available on its microbiological characterization.
In the present study raw and cultured donkey milk samples were analyzed in order to characterize lactic acid bacteria and yeast populations. Both lactic acid bacteria and yeast isolated strains were identified using a combination of different molecular techniques and results were confirmed by sequence analysis of the 16S rRNA gene and ITS regions, respectively. Furthermore, technological traits of the strains were investigated.
The results indicated that dominant lactic acid bacteria strains were identified mainly as Lactococcus lactis subsp. lactis/cremoris and Lactobacillus paracasei. The strains showed both good coagulant and acidifying activities and 50% of them was able to grow in presence of 5.0 g/l of lysozyme. Among isolated yeast strains, the 70% was ascribed to the Kluyveromyces lactis species.
The results of present study indicated that Ragusana donkey milk is an interesting source of lactic acid bacteria and yeast strains of technological interest.
2.Introduction
3. Materials and Methods
3.1 Sampling
3.2 Microbiological analyses
3.3 Microbial isolation
3.4 Genotypic identification of isolates
3.5 Sequence analysis
3.6 Technological characterization of isolated strains
3.7 Statistical analyses
4. Results and Discussions
4.1 Microbiological analysis
4.2 Genotypic identification of isolates
4.3 Yeast identification
4.4 Sequence analysis
4.5 Technological characterization of LAB and yeast strains
5.Conclusion
6.Acknowledgment
5.References
Keywords
LAB; Yeasts; Strain Typing; 16S rDNA; ITS Regions.
Introduction
The interest for donkey milk (DM) is recently growing in terms of cultural and economical importance thanks to its functional proprieties and nutritional value. In Italy, the number of farms breeding donkeys has greatly increased and the most common breeds are Ragusana (2932 livestock), Amiata (2209 livestock), and Sarda (1922 livestock) [1]. DM is considered, among milks from nonbovine mammals, the most suitable for human consumption and its chemical composition is regarded as most similar to human milk [2]. In particular, for its protein profiles DM is recognized as a valid substitute for hypoallergenic formulas for children with both IgE-mediated and non-IgE mediated cow’s milk protein allergy [3, 4]. Furthermore, the low fat content and the high level of polyunsaturated fatty acids, makes the DM recommended for elderly and convalescent consumers [5, 6]. Moreover the high lactose content (60-65 g/l) is relevant for absorption of calcium, for preventing intestine infections and for extending its consumption among kids [7]. Several studies have focused on the content of functional components (bioactive compounds) and antimicrobial substances, such as lysozyme and lactoferrin. In particular, lysozyme plays a significant role in the intestinal immune response [8] and against several undesirable microorganisms.
Although in recent years a growing interest for microbial population of DM has been paid, it has mainly focused onto microbial safety issue.
The aim of the present study was to identify and characterize the dominant lactic acid bacteria (LAB) and yeast population of Ragusana DM through a combination of different molecular techniques, in order to study its microbial heritage and to select strains of technological interest.
Materials and Methods
Milk was obtained from the a farm located in Catania, Sicily (Italy). Milk was collected by mechanical milking, from Ragusana breed donkeys, at middle lactation stage [8].
Nine raw milk samples (RM), 300 ml each, were collected in three consecutive weeks. Samples, transported into the laboratory of Di3A, were split into two aliquots: one was immediately subjected to microbiological analysis, the other one was incubated at 37°C for 24 h, obtaining the cultured milk (CM) samples.
DM samples were serially diluted into quarter-strength Ringer Solution (Oxoid, Ltd, Basingstoke, UK) and plated on: Plate Count Agar (PCA) medium (Oxoid) plus cycloheximide (4 mg/l) (Fluka, Milan, Italy), incubated at 30°C for 48 h, for aerobic mesophilic bacteria; Man Rogosa and Sharpe (MRS) medium (pH 5.6) plus cycloheximide (100 mg/l), anaerobically (using Anaerogen kit) incubated at 32°C for 24-48 h, for lactobacilli; LM17 agar medium, plus cycloheximide (100 mg/l), anaerobically incubated at 32°C for 24-48 h, for lactococci & streptococci; Sabouraud Dextrose Agar (SDA), plus chloramphenicol (100 mg/l) (Fluka), incubated at 25°C for 24-48 h, for yeasts; Violet Red Bile Glucose Agar (VRBGA), incubated at 37°C for 24-48 h, for Enterobacteriaceae. Microbial counts were performed in triplicate for each sample and results were reported as log10 cfu/ml.
In order to isolate the dominant LAB and yeast strains, from both RM and CM samples, 10 colonies were randomly selected from the highest dilution plates of MRS, LM17, and SDA media and purified by re-streaking three times on the same media.
Thirty-two isolates were supposed as LAB, based on Gram reaction, motility, catalase activity and no spore formation. The strains were stored in liquid cultures in 20% glycerol at -20°C.
From SDA plates 50 isolates were microscopically selected for cell morphology and strains were maintained at 4°C in SDA medium slants.
For identification and typing of LAB strains 85 LAB reference strains were used. In details, 43 LAB isolated from Grana Padano cheese (GP), belonging to the Department of Food Science of University of Parma (UPR), 33 LAB reference strains and 9 type strains from BCCM/LMG, Ghent (Belgium) and from DSMZ, Braunschweig Germany) were considered (supplementary Table 1).
All reference/type strains were kept in MRS or in M17 broth containing 20% (v/v) of glycerol at -80°C. The reference/type strains were recovered by two overnight sub-culturing at 32°C. In details, strains belonging to Enterococcus faecalis, Enterococcus faecium, Lactobacillus delbrueckii, Lactobacillus fermentum, Lactobacillus helveticus species were cultivated in MRS at 37°C; strains belonging to Lactobacillus casei, Lactobacillus paraplantarum, Lactobacillus plantarum, and in MRS at 30°C; in LM17 at 30°C and in LM17 at 37°C in anaerobic conditions.
Genomic DNA of LAB strains was extracted from overnight cultures (approximately 8 log cfu/ml). Cultures were centrifuged at 12,000 x g for 5 min (Eppendorf 5810R, Hamburg, Germany), the pellets were suspended in TE buffer (1.0 M Tris-HCl, 0.5 M EDTA, pH 8.0) and then a phenol/ chloroform procedure was carried out [9]. Strain identification was performed by RFLP of amplified tRNAAla-23S rDNA [10]. Briefly, Intergenic Spacer Region (ISR) sequences were amplified using tRNAAla and 23S/p10 primers (Ebesberg, Germany) and digested with HindIII, HinfI and TaqI enzymes [10]. Two additional widely used band profile-based fingerprinting methods, such as the P1 RAPD and (GTG)5 repetitive extragenic palindromic PCR (rep-PCR), were applied [10].
The yeast isolates were identified by PCR/ RFLP of the internal transcribed spacer (ITS) regions [11]. In details, strains were overnight grown in YPD medium (g/l distilled water: yeast extract, 10; peptone, 10; dextrose, 20) (Oxoid) at 28°C under vigorous agitation. DNA was extracted according to Platania et al. [12]. Purified DNA was suspended in a solution containing MyTaq™ Mix (Bioline, London, UK). The rDNA ITS regions were amplified by the ITS1 (5’-TCCGTAGGTGAACCTGCGG- 3’) and ITS4 (5’-TCCTCCGCTTATTGATATGC-3’) primers, in a Personal Thermocycler (Whatman Biometra, Germany), programmed as follows: 94°C for 3 min, 30 cycles of 94°C for 30 s, 55°C for 30 s, 72°C for 1 min, and 72°C for 5min. Aliquots of the PCR-amplified products (5-10 μl) were separately digested with HhaI, HaeIII, HinfI, MspI and RsaI enzymes (New England BioLabs, Beverly, MA) in a final volume of 20 μl. Restriction fragments were quantified in a 2.0% Agarose D1 LE (Conda, Torrejón de Ardoz, Madrid, Spain) gel containing GelRed Nucleic Acid Gel Stain (Biotium Inc., Hayward, CA).
To confirm the species attribution the 16S rRNA gene and the ITS regions were sequenced [13, 14]. The sequences were compared to those present in public data libraries GenBank and EZ-Taxon using the BLAST search program (www.ncbi.nlm.nih.gov/BLAST/; ezgenome.ezbiocloud.net/ezg_BLAST).
LAB strains were overnight pre-cultured at 32°C in MRS broth for technological characterization. The ability to grow at 4, 15, and 45°C was monitored in MRS broth by optical density readings at 620 nm after 5, 3, and 2 days, respectively. The ability to acidify sterile skim milk powder (reconstituted at 100 g/l) containing 0.1% of Yeast Extract (Oxoid, Italy) was determined by a pH-meter (Eutech Instruments, XSPH 510, The Netherlands) after 8 h of incubation at 32°C. The coagulating activity was visually evaluated by monitoring the appearance of a coagulum on the inner site of a glass tube containing 10% (w/v) of skim milk powder. The ability to grow at different lysozyme concentrations was determined in MRS medium containing lysozyme at 1.5, 2.5 or 5.0 g/l (Sigma Chemical Co., Milan, Italy) after 24 h of incubation at 32°C, in anaerobic conditions.
Regarding yeast, the vegetative reproduction (by multilateral, bipolar or unipolar budding, by fission, by forming filaments) was observed after incubation in Yeast Dextrose Peptone and Malt Yeast Glucose Peptone media [15]. Growth at 25, 32, 37, and 42°C was visually determined after 2 and 4 days of incubation.
All microbial counts were reported as average values and Standard Deviations. Statistical ANOVA and Duncan tests were performed using XLSTAT PRO 5.7 (Addinsoft, New York, USA). Statistical ANOVA analysis was carried out to determine differences among samples for each microbial group.
LAB genotyping data were analysed exporting TIFF files from the Odyssey scanner into the pattern analysis software package BioNumerics (package version 5.1; Applied Maths, BVBA, Sint- Martens-Latem, Belgium). Using the “composite data set” of the software, isolate rDNA-RFLP band profiles, obtained by HindIII, HinfI and TaqI digestions, were combined with profiles of the internal database comprising band profiles from the 85 reference strains obtaining a unique dendrogram. The identification of LAB strains was determined by strain cluster association and a single dendrogram was obtained also from RAPD and rep-PCR fingerprinting. DNA patterns were analysed through the unpaired group method with arithmetic average (UPGMA). Calculation of similarity of all fingerprinting profiles was based on Dice correlation coefficient.
The mean microbial counts (log cfu/ml), detected both in raw milk (RM) and in cultured milk (CM) samples, at the three consecutive
weeks, and after incubation at 37°C for 24h, respectively, are reported in Figure 1. Mesophilic aerobic bacteria exhibited an average value of approximately 3.3 log cfu/ml in RM samples, in compliance to the Commission Regulation (EC) No 1662 [16], and in agreement with previous studies [17, 18]. These results confirm that the mesophilic aerobic bacteria values in DM are lower than those reported for bovine or sheep milk [19, 20] (Conte et al., 2004; Cavallarin et al., 2015), probably for the presence of antimicrobial substance [21, 22].
Lactobacilli counts showed significant differences with an average value of about 2.0 log cfu/ml in RM samples, and 6.0 log cfu/ ml in CM samples (Figure 1). Lactococci & streptococci counts were always higher than lactobacilli, according to Carminati et al.[23]. These results could be correlated to the higher resistance of thermophilic cocci-LAB to lysozyme [24]. The final pH value, in CM samples, dropped from 7.12+0.08 to 5.28+0.03, confirming previous reports [17, 20].
The mean values of yeast counts were 2.0 and 4.5 log cfu/ml, in RM and CM samples, respectively (Figure 1), in agreement with Coppola et al. [21] and in contrast to Šarić et al. [25] that did not find yeast in any of analyzed DM samples. The higher yeast density in DM, compared to those detected in cow’s milk, confirmed that lysozyme does not affect the yeast growth [17, 21].
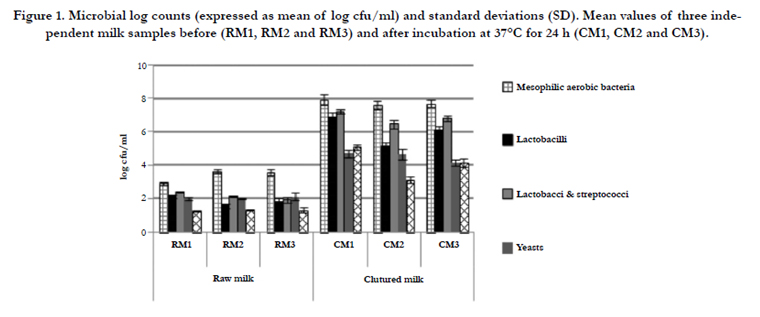
Figure 1. Microbial log counts (expressed as mean of log cfu/ml) and standard deviations (SD). Mean values of three independent milk samples before (RM1, RM2 and RM3) and after incubation at 37°C for 24 h (CM1, CM2 and CM3).
A total of 32 LAB strains were selected from both MRS and LM17 media, both from RM and CM samples.
In this work, the affiliation level of isolated LAB species was assessed using the tRNAAla-23S rDNA-RFLP fingerprinting method [10]. The RFLP profiles from the amplified ISR portion were clustered with the set of “reference database” LAB dairy strains, using the BioNumerics software, obtaining a unique dendrogram (Figure 2). Four genotype clusters were clearly formed (cluster 1, 2, 3 and 4) (Figure 2).
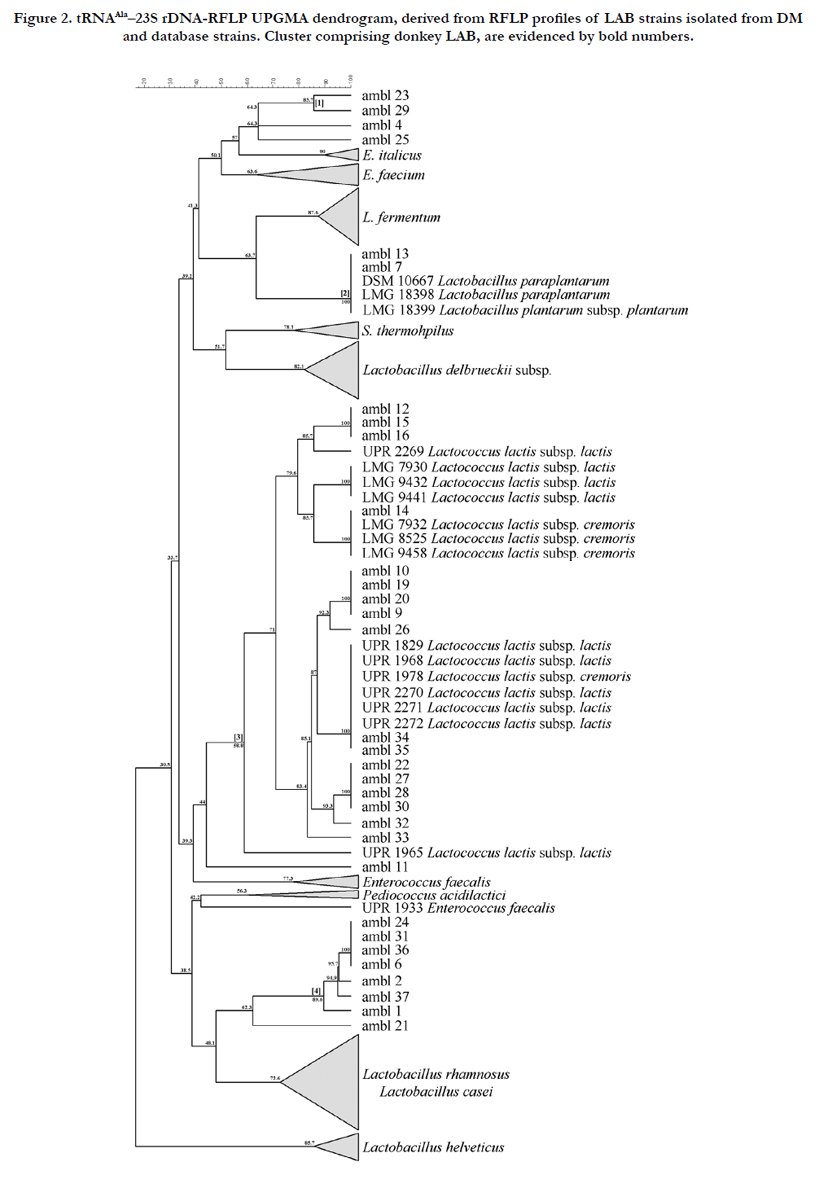

Figure 2. tRNAAla–23S rDNA-RFLP UPGMA dendrogram, derived from RFLP profiles of LAB strains isolated from DM and database strains. Cluster comprising donkey LAB, are evidenced by bold numbers.
Four LAB strains (AMBL23, -29, -4 and -25), at the similarity level of 64.3%, were grouped in cluster 1, without any association with the internal database strains. Even if they belong to the biggest Enterococcus dendrogram branch (50.1% similarity level), they showed higher similarity to E. italicus reference strain rather than to E. faecium.
The strains AMBL13 and -7 were clustered together with L. plantarum into cluster 2. Seventeen isolates were included in cluster 3, together with Lc. lactis subsp. lactis/cremoris GP-type strains, with a similarity level of 59%, while the AMBL11 strain could be considered in the same cluster. Eight strains formed the independent cluster 4, which did not match to any reference strains. In order to confirm the species affiliation, 13 strains, representing each clusters, were subjected to the 16S rRNA sequencing (Table 1). Results revealed that all strains from cluster 1 were identified as E. faecium, while the AMBL13 (cluster 2), was identified as L. plantarum subsp. plantarum. Strains of cluster 3, including the AMBL11, were identified as Lc. lactis subsp. lactis, and strains of cluster 4 as L. paracasei. In order to overcome the sequence identification limits between L. casei and L. plantarum groups, specific and multiplex PCRs were performed. The results mirrored the sequencing data (data not shown). Cluster 4 remained distant from the L. casei/rhamnosus reference strain cluster, at the similarity value of 48%.
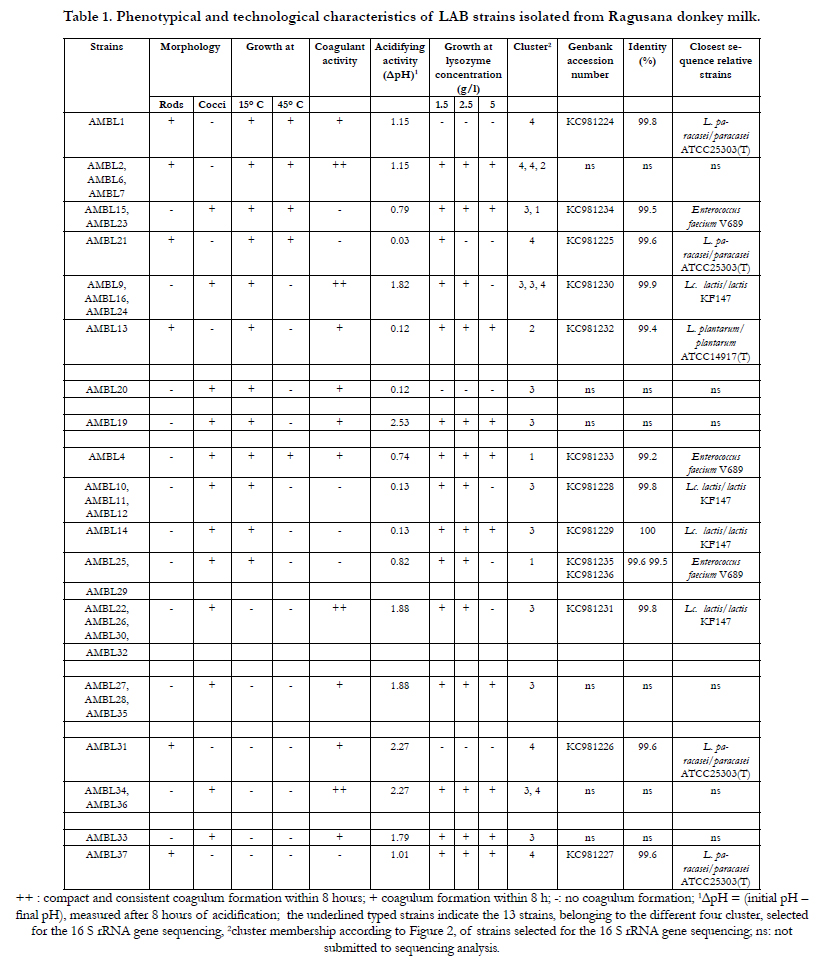

Table 1. Phenotypical and technological characteristics of LAB strains isolated from Ragusana donkey milk.
In order to find out about interspecies diversity and single species biotype composition for each LAB cluster, RAPD-PCR and (GTG)5 rep-PCR were carried out. RAPD-PCR is the most widely used method for LAB characterization in dairy products [13, 26] and the Rep-PCR is often applied for typing biotypes belonging to the same species [27, 28]. P1 RAPD and (GTG)5 rep-PCR fingerprinting profiles of the Lc. lactis strains were combined and analysed with profiles of the Lc. lactis reference internal strains and five clusters were obtained, revealing that most Lc. lactis strains (fifteen) were grouped in two clusters (2 and 5) distinct from reference internal strains and from the GP strains (Figure 3). Moreover, the AMBL9, -10, -11 strains were grouped in cluster 3 together with the reference strain Lc. lactis subsp. cremoris LMG9458 (Figure 3). Even if this approach did not allow to discriminate between the subspecies lactis/cremoris, it revealed a high degree of intra-species heterogeneity. Within Lc. lactis the two genotype discrimination is still difficult and it has reported that strains of subspecies lactis often show a cremoris genotype [29], and vice-versa [30]. Hence, the phenotypic and genetic relationships between and within the subspecies of Lc. lactis remain unclear.
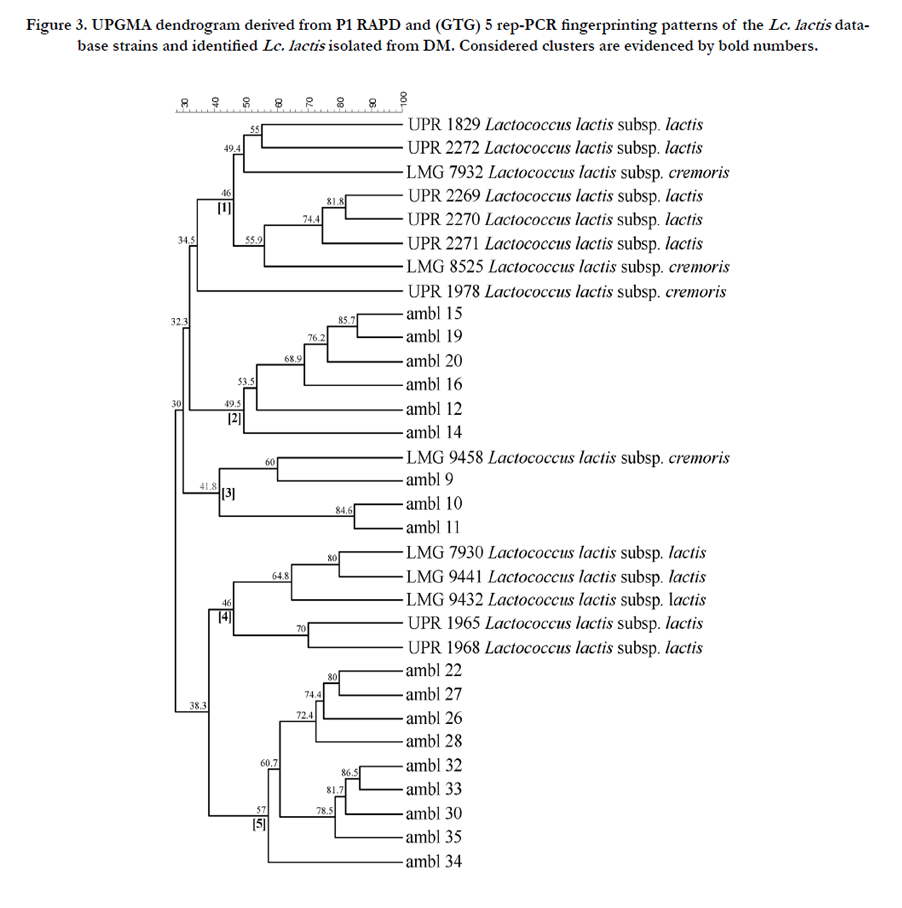
Figure 3. UPGMA dendrogram derived from P1 RAPD and (GTG) 5 rep-PCR fingerprinting patterns of the Lc. lactis database strains and identified Lc. lactis isolated from DM. Considered clusters are evidenced by bold numbers.
Results of present study are inconsistent with previous identification of LAB in DM, reporting the presence of mesophilic lactobacilli as L. paracasei, L. brevis, L. salivarius, and L. plantarum [31]. The biotype analysis of the eight L. casei strains demonstrates that, except for the AMBL21 strain, the similarity with the reference strains was less than 38% (Figure 4).

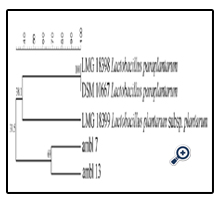
Figure 4. UPGMA dendrogram derived from P1 RAPD and (GTG)5 rep-PCR fingerprinting patterns of the L. casei database strains and identified L. paracasei isolated from DM.
LAB strains belonging to Lactobacillus genus have been divided in two groups, one attributed to L. casei, and one to L. plantarum/L. paraplantarum. Regarding the L. casei group low similarity level (48%) were detected. Although the isolation of L. plantarum from DM has been recently reported [31] in the present study it was not allowed the discrimination among L. plantarum, L. plantarum subsp. plantarum and L. pentosus species, as previously reported [10, 32].
The four strains identified as E. faecium (Figure 5) showed a different biotype affiliation, with a similarity of 52.7% to LMG strains, except the AMBL4 strain that remained distant (similarity of 38.6%). The last group of LAB belonged to E. italicus species, firstly isolated from artisanal Italian cheeses [33]. The presence of enterococci in dairy products has long been reported. Carminati et al. [23] correlated the presence of E. faecalis isolates in DM to their high resistance to lysozyme.
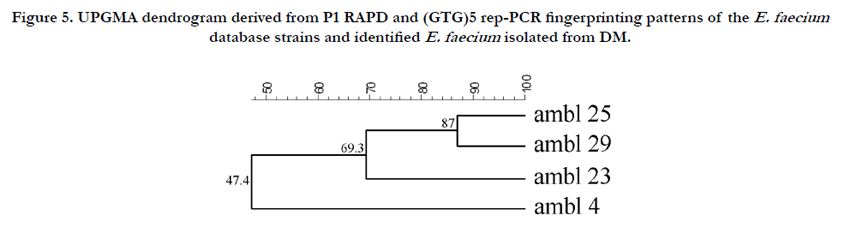
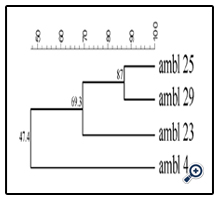
Figure 5. UPGMA dendrogram derived from P1 RAPD and (GTG)5 rep-PCR fingerprinting patterns of the E. faecium database strains and identified E. faecium isolated from DM.
Although the monitoring of fungi of technological interest for the cheese-making industry is considered relevant, few studies have focused on the composition of fungal communities in milk [34, 35].
In the present study 50 yeast strains were identified (Table 2). The PCR amplification of the 5.8 ITS regions yielded four amplicons, ranging from 530 to 740 bp, and subsequent digestion with HhaI, HaeIII, and HinfI revealed 4 presumptive different species. The HinfI enzyme discriminated between Kluyveromyces marxianus and Kluyveromyces lactis [36]. In addition, the RsaI enzyme ascribed 36 yeast strains to Kluyveromyces lactis species. The enzyme MspI, generating a fragment from 740 to 300 bp, was used to discriminate Candida albicans from other species [37]. The species K.lactis (cluster 1), Candida albicans (cluster 2) and Cryptococcus curvatus (cluster 3) were identified by comparing the restriction fragments with those previously described [36-38]. Strains belonging to the cluster 4 showed a RFLP profile which did not match to any reported data. The ITS region sequencing confirmed the identification based on restriction analysis of the 5.8 ITS region, except for cluster 2 which was ascribed to the species Trichosporon moniliforme. In the case of cluster 4, the ITS region sequencing ascribed the 7 strains within the species Candida parapsilosis (98% of similarity). Yeasts species detected in raw milk include K. marxianus, K. lactis, Rhodotorula mucilaginosa, Debaryomyces hansenii, Geotrichum candidum, Geotrichum catenulate, Pichia fermentans, Candida sake, Candida parapsilosis, Candida inconspicua, Trichosporon cutaneum, Trichosporon lactis, Cryptococcus curvatus, Cryptococcus carnescens and Cryptococcus victoriae [35]. In the present study the 72% of yeast strains was assigned to the K. lactis species, known to convert lactose into lactic acid [39, 40] and to play a positive role in development of aroma, texture, digestibility, and/ or determining the growth of other microorganisms [41]. The remaining yeast strains were identified as Cry. curvatus, T. moniliforme and C. parapsilosis. The three strains of cluster 2, identified as T. moniliforme, were firstly ascribed to the C. albicans, because the MspI enzyme cuts is in the high homology fragment within the internal sequence of the ITS region. Trichosporon genus has been frequently isolated from milk and dairy products [42]. The species C. parapsilosis is one of the most frequently isolated yeast in raw ewe milk [35].
Results of the present work showed that the dominant bacterial species were identified as Lc. lactis/cremoris (50%), L. paracasei (25%), E. faecium (12%), and L. plantarum (6%), whereas Carminati et al. [23] found Streptococcus macedonicus (45%), E. faecalis (28%), E. faecium (16%). In the present study the 72% of isolated yeast strains was ascribed to the lactose fermenting species K.lactis.
The sequences of the LAB 16S rRNA gene and yeast ITS regions were deposited in the GenBank database. The accession numbers of the 13 selected bacterial strains are reported in Table 1 and those related to the 4 yeast strains in Table 2.
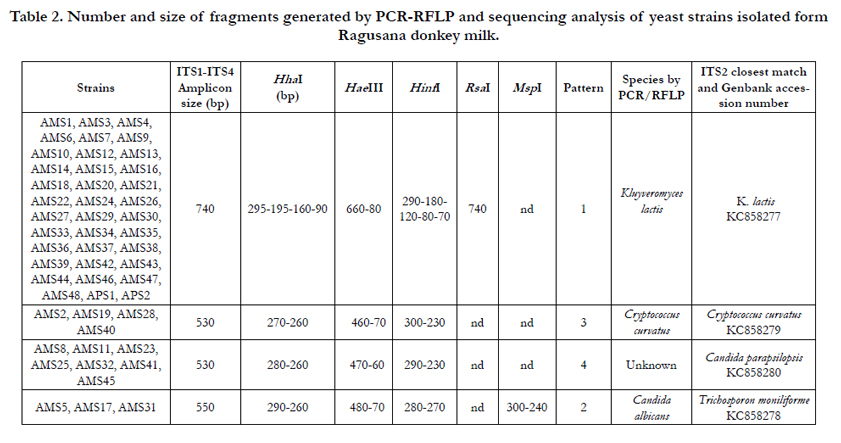
Table 2. Number and size of fragments generated by PCR-RFLP and sequencing analysis of yeast strains isolated form Ragusana donkey milk.
The results of characterization of the 32 LAB strains confirmed their high heterogeneity (Table 1) and allowed to distinguish 18 different phenotypic clusters. The 75% of the strains showed a coccal morphology and the 80% showed a homofermentative metabolism (data not shown). No strain exhibited growth at 4°C, whereas the 62% grew at 15°C, and 40% of them also at 45°C. Twenty-two strains (68%) showed good coagulant activity, and twelve of them (54%) showed a very good coagulant activity, with a formation of a compact coagulum within 8 h (Table 1).
Regarding acidifying activity results showed that: four strains (13%), belonging both to L. casei/paracasei and Lc. lactis species, exhibited a very good acidifying activity (ΔpH > 2.0); sixteen strains (50%), ten identified as Lc. lactis, five as L. paracasei and one as L. plantarum, exhibited a ΔpH value between 1.0 and 2.0; five strains (16%), four identified as E. faecium showed a ΔpH value between 0.5 and 1.0. The highest acidification activity was revealed by Lb. paracasei strains. The most Lc. lactis strains exhibited a ΔpH> 1.0.
Among facultative heterofermentative LAB the importance of L. paracasei species in dairy field is linked to its role as non-starter LAB and in flavour development during cheese ripening [43].
Even though LAB cocci have generally been considered more resistant to lysozyme than lactobacilli, results of the present study highlighted that the 60% of lactobacilli and the 45% coccus shaped LAB showed growth at the highest lysozyme concentration tested (Table 1), confirming that resistance to lysozyme is a strain specific trait [24].
Results of technological characterization of the 50 yeast strains are showed in the Table 2. No pigment production was detected, while three different morphological types were observed. All strains grew at 25, 32 and 37°C, and 45 (90%) grew also at 42°C (data not shown).
Conclusion
In the present work the cultivable microbial community of Ragusana DM has been investigated and results revealed a high diversity among both LAB and yeasts strains. The dominant LAB strains were identified as Lc. lactis subsp. lactis/cremoris and L. paracasei and the K. lactis species dominated within the yeast population. Most of the LAB strains exhibited good technological performances, demonstrating that Ragusana DM is an interesting source for selecting starter cultures for dairy industry.
Acknowledgment
We would like to thank the ASILAT farm (Giarre, Catania) for providing donkey milk samples.
References
- AIA, Associazione Italiana Allevatori, Equidi per regione. (2016) www.anagrafeequidi.it/index.php?id=217. Accessed on 10 March 2016.
- Salimei E, Fantuz F, Simoni A, Varisco G, Chiari C (2004) Ass’s milking machine: first results on residual milk. In: Proceedings of 6th congress new findings in equine practices, Campobasso, Italy pp 93–98.
- Muraro A, Dreborg S, Halken S, Høst A, Niggemann B, Aalberse R, et al. (2004) Dietary prevention of allergic diseases in infants and small children. Part I: Immunologic background and criteria for hypoallergenicity. Pediatr Allergy Immunol 15(2): 103–111.
- Restani P, Ballabio C, Di Lorenzo C, Tripodi S, Fiocchi A (2009) Molecular aspects of milk allergens and their role in clinical events. Anal Bioanal Chem 395(1): 47–56.
- Fantuz F, Ferraro S, Todini L, Piloni R, Mariani P, et al. (2012) Donkey milk concentration of calcium, phosphorus, potassium, sodium and magnesium. Int Dairy J 24(2): 143-145.
- Salimei E, Fantuz F (2012) Equid milk for human consumption. Int Dairy J 24(2): 130–142.
- Schaafsma G (2003) Nutritional significance of lactose and lactose derivates. In H Roginski, JW, Fuqua, P F, Fox. (Eds) Encyclopedia of Dairy Science. London: Academic Press 3: 1835-1842.
- Tidona F, Sekse C, Criscione A, Jacobsen M, Bordonaro S, et al. (2011) Antimicrobial effect of donkeys’ milk digested in vitro with human gastrointestinal enzymes. Int Dairy J 21(3): 158–165.
- de Los Reyes-Gavilán CG, Limsowtin GK, Tailliez P, Sechaud L, Accolas JP (1992) A Lactobacillus helveticus-specific DNA probe detects restriction fragment length polymorphisms in this species. Appl Environ Microbiol 58(10): 3429–3432.
- Mancini A, Lazzi C, Bernini V, Neviani E, Gatti M (2012) Identification of dairy lactic acid bacteria by tRNAAla-23S rDNA-RFLP. J Microbiol Meth 91(3): 380–390.
- Esteve-Zarzoso B, Belloch C, Uruburu F, Querol A (1999) Identification of yeasts by RFLP analysis of the 5.8S rRNA gene and the two ribosomal internal transcribed spacers. Int J System Bacteriol 49(1): 329–337.
- Platania C, Restuccia C, Muccilli S, Cirvilleri G (2012) Efficacy of killer yeasts in the biological control of Penicillium digitatum Tarocco orange fruits (Citrus sinensis). Food Microbiol 30(1): 219–225.
- Giraffa G, Rossetti L (2004) Monitoring of the bacterial composition of dairy starter cultures by RAPD-PCR. FEMS Microbiol Lett 237(1): 133–138.
- Leaw SN, Chang HC, Sun HF, Barton R, Bouchara JP, Chang TC (2006) Identification of medically important yeast species by sequence analysis of the internal transcribed spacer regions. J Clin Microbiol 44(3): 693–699.
- Barnett JA, Payne RW, Yarrow D (1990) Yeasts: Characteristics and Identification.( 2nd edn), Cambridge University Press, Cambridge.
- European Union (2006) Commission Regulation (EC) No 1662/2006 of 6 November 2006 amending Regulation (EC) No 853/2004 of the European Parliament and of the Council laying down specific hygiene rules for food of animal origin. Official Journal of the European Union L320: 1-10.
- Zhang X-Y, Zhao L, Jiang L, Dong M-L, Ren FZ (2008) The antimicrobial activity of donkey milk and its microflora changes during storage. Food Control 19(12): 1191-1195.
- Alabiso A, Giosuè C, Alicata ML, Mazza F, Iannolino G (2009) The effects of different milking intervals and milking times per day in jennet milk production. Animal 3(4): 543–547.
- Conte F, Scatassa ML, Monsù G, Lo Verde V, Finocchiaro A, et al. (2004) Monitoring of safety and quality of donkey’s milk. In Proceeding of the International Conference Veterinary Public Health and Food Safety. Towards a risk based chain control. 58–59.
- Cavallarin L, Giribaldi M, Soto-Del Rio MD, Valle E, Barbarino G, et al. (2015) A survey on the milk chimical and microbiological quality in dairy donkey farms located in NorthWestern Italy. Food Control 50: 230-235.
- Coppola R, Salimei E, Succi M, Sorrentino E, Nanni M, Ranieri P (2002) Behaviour of Lactobacillus rhamnosus strains in ass’s milk. Ann Microbiol 52: 55–60.
- Chiavari C, Coloretti F, Nanni M, Sorrentino E, Grazia L (2005) Use of donkey’s milk for a fermented beverage with lactobacilli. Le Lait, 85(6): 481- 490.
- Carminati D, Tidona F, Fornasari ME, Rossetti L, Meucci A, Giraffa G (2014) Biotyping of cultivable lactic acid bacteria isolated from donkey milk. Lett Appl Microbiol 59(3): 299-305.
- Neviani E, Carminati D, Veaux M, Hermier I, Giraffa G (1991) Characterization of Lactobacillus helveticus strains resistant to lysozyme. Le Lait 71(1): 65-73.
- Saric LC, Saric BM, Mandic AI, Torbica AM, Tomić JM, et al. (2012) Antibacterial properties of domestic Balkan donkeys’ milk. Int Dairy J 25(2):142–146.
- Monfredini L, Settanni L, Poznanski E, Cavazza A, Franciosi E (2012) The spatial distribution of bacteria in Grana-cheese during ripening. Syst Appl Microbiol 35(1): 54–63.
- Bove CG, De Dea Lindner L, Lazzi C, Gatti M, Neviani E (2011) Evaluation of genetic polymorphism among Lactobacillus rhamnosus non-starter Parmigiano Reggiano cheese strains. Int J Food Microbiol 144(3): 569–572.
- Coudeyras S, Marchandin H, Fajon C, Forestier C (2008) Taxonomic and strain-specific identification of the probiotic strain Lactobacillus rhamnosus 35 within the Lactobacillus casei group. Appl Environ Microbiol 74(9): 2679–2689.
- Jarvis AW, Jarvis BDW (1981) DNA homology among lactic streptococci. Appl Environ Microbiol 41: 77–83.
- Tanigawa K, Kawabata H, Watanabe K (2010) Identification and typing of Lactococcus lactis by matrix-assisted laser desorption ionization–time of flight mass spectrometry. Appl Environ Microbiol 76(12): 4055–4062.
- Murua A, Todorov SD, Vieira ADS, Martinez RCR, Cenić A, Franco BDGM (2013) Isolation and identification of bacteriocinogenic strain of Lactobacillus plantarum with potential beneficial properties from donkey milk. J Appl Microbiol 114(6): 1793-1809.
- Rachman CN, Kabadjova P, Prevost H, Dousset X (2003) Identification of Lactobacillus alimentarius and Lactobacillus farciminis with 16S-23S rDNA intergenic spacer region polymorphism and PCR amplification using species- specific oligonucleotide. J Appl Microbiol 95(6): 1207–1216.
- Fortina MG, Ricci G, Borgo F, Manachini PL (2007) Rapid identification of Enterococcus italicus by PCR with primers targeted to 16S rRNA gene. Lett Appl Microbiol 44(4): 443–446.
- Cocolin L, Aggio D, Manzano M, Cantoni C, Comi G (2002) An application of PCR–DGGE analysis to profile the yeast population in raw milk. Int Dairy J 12(5): 407–411.
- Delavenne E, Mounier J, Asmani K, Jany JL, Barbier G, Le Blay G (2011) Fungal diversity in cow, goat and ewe milk. Int J Food Microbiol 51(2): 247–251.
- Belloch C, Barrio E, García MD, Querol A (1998) Phylogenetic reconstruction of the yeast genus Kluyveromyces: restriction map analysis of the 5.8S rRNA gene and the two ribosomal internal transcribed spacers. Syst Appl Microbiol 21(2): 266–273.
- Mirhendi SH, Makimura K, Khoramizadeh M, Yamaguchi H (2006) A oneenzyme PCR-RFLP assay for identification of six medically important Candida species. Jpn J Med Mycol 47(3): 225–229.
- Bockelmann W, Heller M, Heller K (2008) Identification of yeasts of dairy origin by amplified ribosomal DNA restriction analysis (ARDRA). Int DairyJ 18(10): 1066–1071.
- Chang YD, Dickson RC (1988) Primary structure of the lactose permease gene from the yeast Kluyveromyces lactis. J Biol Chem 263(32): 16696– 16703.
- Poch O, L’Hote H, Dallery V, Debeaux F, Fleer R, Sodoyer R (1992) Sequence of the Kluyveromyces lactis β-galactosidase: comparison prokaryotic enzymes and secondary structure analysis. Gene 118(1): 55–63.
- Law BA (2001) Controlled and accelerated cheese ripening: the research base for new technologies. Int Dairy J 11(4): 383–398.
- Corbo MR, Lanciotti R, Albenzio M, Sinigaglia M (2001) Occurrence and characterization of yeasts isolated from milks and dairy products of Apulia region. Int J Food Microbiol 69(1): 147–152.
- El Soda M, Madkor SA, Tong PS (2000) Adjunct cultures: recent developments and potential significance to the cheese industry. J Dairy Sci 83(4): 609–619.





