Effects of 5-Azacytidine on Differentiation of Ovine Mesenchymal Stem Cells
Soltani L1, Rahmani H1, Joupari MD2*, Ghaneialvar H3, Amirhossein Mahdavi1, Shamsara M2
1 Department of Animal Science, Faculty of Agriculture, Isfahan University of Technology, Isfahan, Iran.
2 Department of Animal and Marine Biotechnology, National Institute of Genetic Engineering and Biotechnology (NIGEB), Tehran, Iran.
3 Department of Clinical Biochemistry, Faculty of Medical Science, Tarbiat Modares University, Tehran, Iran.
*Corresponding Author
Morteza Daliri Joupari,
Department of Animal and Marine Biotechnology,
National Institute of Genetic Engineering and Biotechnology (NIGEB),
Tehran, Iran.
Tel: +(98)9122609195
Email: Daliri@nigeb.ac.ir
Article Type: Research Article
Received: April 25, 2015; Accepted: May 27, 2015; Published: June 01, 2015
Citation: Joupari MD et al., (2015) Effects Of 5-Azacytidine On Differentiation Of Ovine Mesenchymal Stem Cells. Int J Stem Cell Res Transplant 03(2), 96-100. doi: dx.doi.org/10.19070/2328-3548-1500016
Copyright: Joupari MD© 2015. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
Background: Mesenchymal stem cells (MSCs) have potential of self-renewal and differentiation into many cell types and can be used for cells therapy. Addition of epigenetic drugs can also convert these cells into cardiomyocytes.
Objective: The aim of this study was to evaluate the effect of 5-azacytidine on differentiation of sheep fetal bone marrow MSCs into cardiomyocytes.
Materials and Methods: Isolated bone sheep fetal marrow MSCs was cultured in DMEM: F12 medium. Passaged-3 cells were examined for their differentiation capacity into osteocytes and adipose cells. Surface antigens of isolated cells were analyzed using flow cytometry. Cells after third passage treated with 5 μM 5-azacytidine for 48h followed by medium without 5-azacytidine for twenty-one days. Expression of cardiac alpha-actinin and Troponin I (cTnI) were analyzed by immunohistochemistry.
Results: MSCs expressed CD44, CD166 and weak expression of CD34 and CD45, apart from this expression a very weak expression of CD105 was also observed. Oil red for adipose cells and alizarin red staining for osteocytes indicated that the cells from all studied groups maintained the differentiation potential into adipose and osteocytes cells. Expression of Gata4, connexin 43, Atrial Natriuretic Peptide (ANP) was also detected by reverse transcriptase polymerase chain reaction and weak expression of troponinI. Sarcomeric alpha-actinin and cTnI were also partially detected by immunohistochemistry on these cells.
Conclusion: This result indicated that addition of 5-azacytidine into culture medium could be effective on MSCs differentiation into cardiomyocytes.
2.Introduction
3.Materials and Methods
3.1.Chemicals
3.2.Isolation, cell culture, and phenotypic characterization
3.3.Differentiation of cardiomyocyte-like cells
3.4.Immunohistochemistry analysis
4.Results
5.Discussion
6.Conclusion
7.Acknowledgement
8.References
Keywords
5-Azacytidine; Mesenchymal Stem Cells; Sheep Fetal; Cardiomyocytes Differentiation.
Introduction
Myocardial infarction among the major diseases strongly influenced the Longevity and quality of life. Human adult cardiomyocytes has a limited capacity for regeneration after myocardial infarction, the irreversible loss of cardiomyocytes, and loss of contraction muscle andincrease proliferation and replacement of fibroblast cells can lead to progressive ventricular remodeling of nonischaemic myocardium, and the ventricular remodelling can result in progressive ventricular dilatation and heart failure [1]. Clinically there is no treatment to regeneration of infarcted myocardium. On this basis, cell therapy is ideal treatment to regenerate the damaged cardiac area.
A lot of studies tried to improve cardiac function after myocardial infarction with cell therapy; namely: using embryonic stem cells [2] and mesenchymal stem cells (MSCs) [3, 4]. MSCs have some advantages than others because they do not form teratoma and will not raise any ethical problems as embryonic stem cells do. Using of MSCs provides basic fundamental of mechanism of fate and function of cell improvement.
Different strategies for differentiation of MSCs into cardiomyocytes can be used. Small molecules such as ascorbic acid, dexamethasone, all-trans retinoic acid and 5-azacytidine can induce differentiation of stem cells into cardiomyocytes [5]. Demethylating epigenetic drugs have gained interest because of their reprograming effects during stem cell differentiation [6].
Makino et al., for the first time used 5-azacytidine in order that differentiation of mouse MSCs, they showed that mouse MSCs can differentiate into beating cells [7]. Addition of 10 μM 5-azacytidine in culture of hMSCs for 28 days could induce differentiation towards cardiomyogenic lineage, whereas same cultural condition for a period of 24h lead to cardiac-like muscle cells [8, 9]. Fukuda et al. showed that eight weeks after treatment with 3 mM 5-azacytidinea cardiomyogenic cell line from murine bone marrow stromal [10].
The better model for studying human pathology than rodent models are domestic animals because immunologically and physiologically they are similar to human [11].
On the basis of research reports there is little information available about the effects of 5-azacytidine on ruminant bone marrow MSCs, we therefore designed our experiment to evaluate the effect of 5-azacytidine (small molecule) on differentiation of sheep fetal MSCs into cardiomyocytes.
Materials and Methods
Except where otherwise indicated, all chemicals were obtained from Sigma-Aldrich (USA).
Ovine fetus (30-35 days) obtained from slaughtered animal at Rock slaughterhouse, Karaj. Fetuses were transported to laboratory in Dulbecco,s PBS on ice. The Bone marrow MSCs were collected from ovine fetus by aspirating femurs and tibias with- DMEM: F12 (3:1) culture media supplemented with 100 IU/ml penicillin, 100 IU/ml streptomycin. Bone marrow aspiration sample was placed on top of ficoll (equal volume) and centrifuged at a speed of 4000 rpm for 30 min at room temperature. Cloudy layer collected and transferred into new tube and washed twice with same medium. The mononuclear cells with concentration of 5×106 cells/ml were cultured in DMEM: F12 supplemented with 5% fetal calf serum (FCS), L-glutamin and penicillin/streptomycin. Cells were incubated at 37°C with 5% CO2. The medium was changed every 3 days after initial plating. After 80% confluency, cells were subcultured using 0.25% trypsin/1 mM EDTA.
Undifferentiated sheep fetal MSCs surface markers were examined by flow cytometry. Sheep fetal MSCs were cultured as described above, cells were centrifuged and pelletwas resuspended in cold DPBS with 108 cells/ml. Cells were incubated with primary antibodies (CD34-FITC; CD44-FITC; CD45-FITC; CD90-FITC and CD166-FITC) followed by secondary conjugated antibody. Isotype-matched antibody (mouse anti-human IgG) was used as negative control, treated cells were incubated at 4°C for 30 min, and the complex was analyzed by flow cytometry.
After three passages of MSCs, cells were cultured in 6-well plates. When cells reached at 80% confluency, they were treated with 5 μM 5-azacytidine for 48h, then 5-azacytidine was deleted from culture medium and again cultured for 21 days, medium was changed every three days. MSCs cultured in complete medium without 5-azacytidine was used as control group.
Total RNA was extracted with AccuZol (Bioneer) from induced and uninduced MSCs. Transcriptional expression of myocardiumspecific Connexin 43, ANP (Atrial Natriuretic Peptide), GATA4, Troponin I was determined by semi-quantitative RT-PCR using the RevertAidTM First Strand cDNA Synthesis Kit (Fermentas, USA) and PCR master mix (Fermentas, USA) according to the manufacturer’s instructions. Transcript levels were standardized to the corresponding sheep GAPDH level. The forward and reverse primers for each PCR set were designed to be located by the Oligo6 software [Connexin 43: For: GTCGTGTCGTTGGTGTCTCTTG,Rve: GCCAGTGGTAGTGTGGTAAGG (114 bp), Troponin I: For: CCGCTCGTCAGCCAACTAC, Rev: GAGTTCCTGCTTCGCAATCTG (124 bp), Gata4: For: GGGCGAGCCAGTGTGTAATG, Rev: GGACCTGCTGGTGTCTTTGATTTG (141bp), ANP: For: ACCGTTTGGAGGACAAGATGC, Rev: TCTCTCTGGGCTGCGTTGAC (147bp), GAPDH: For: ATCGTGGAGGGACTTATGACC; Rev: CGCCAGTAGAAGCAGGGATG (130bp). The thermal profile for PCR was 95 ºCfor 3 min, followed by 40 cycles of 45 sec at 95 ºC, with 1 min annealing intervals (57.8 for GAPDH, Troponin I, Gata4, ANP and 58.2 for Connexin 43) followed by 1 min extension at 72 ºC. Additional 10-min incubation at 72ºC was included after completion of the last cycle. Following the PCR, 5 μl ofthe PCR product was electrophoresed on a 1% agarose geland photographed under UV light.
Induced and uninduced cells were fixed in 4% paraformaldehyde in PBS for 15 minutes, after rinsing with PBS cells were permeabilised in PBS containing 0.2% Triton X-100 for 30 minutes and blocked in blocking buffer containing PBS supplemented with 0.1% BSA and 0. 1% Triton X-100. Primary antibodies were alpha-actinin (A7811, Sigma) and troponin I (cTnI) (Abcam, ab47003). Antibodies were diluted 1:500 for alpha-actinin and 1:300 for troponin I in PBS containing 1% BSA. Cells were incubated in the appropriate antibody solution at 4°C overnight. They were rinsed three times with PBS for 15 minutes each. Cells were incubated with FITC (F9137 Sigma) conjugated secondary antibody diluted 1:400 with PBS for 2 hours at room temperature. Cells were then rinsed three times with PBS. Nuclear staining was performed by 5 min incubation with DAPI (4,6-diamidino- 2-phenylindole). Images were analyzed with a florescent inverted microscope (TE2000; Nikon).
Results
Cultured MSCs grew rapidly and expanded in primary cultures media, non adherent cells were removed while changing the media after 48h. Cells after 5 days incubation showed 80% confluency (Figure 1).
Ovine BMSCs at third passage were evaluated for potential differentiate into adipogenic and osteogenic lineages. Twenty one days after osteogenic induction, BMSCs began to mineralize their matrix. Cultured cells were stained positively with Alizarin Red. BMSCs differentiated cell to adipocytes formed the lipid droplets were also stained with Oil red (Figure 2).
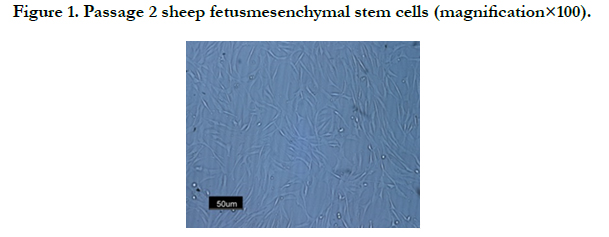
Figure 1. Passage 2 sheep fetusmesenchymal stem cells (magnification×100).
Flow cytometric analysis (Figure 2) showed that the isolated cells at passage 3 expressed the surface markers CD44 (93.48%), CD166 (96.28%) in contrary to the results reported in the human, we also did not get positive expression of CD105 (6.30%). Cells were negative for surface markers of hematopoietic stem cells such as CD34 (6.52%) and CD45 (16.80%).
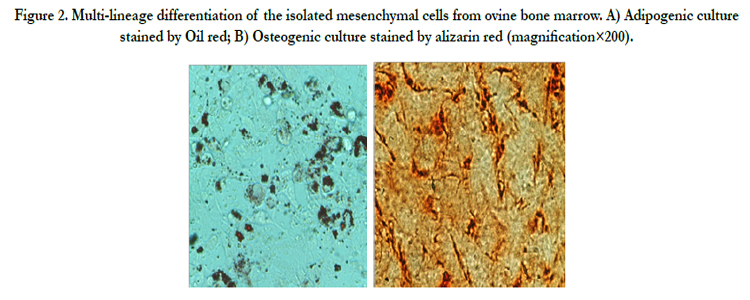
Figure 2. Multi-lineage differentiation of the isolated mesenchymal cells from ovine bone marrow. A) Adipogenic culture stained by Oil red; B) Osteogenic culture stained by alizarin red (magnification×200).
RT-PCR analysis for the expression of cardiomyocyte genes represented that both induced and uninduced cells expressed connexin 43 and GAPDH (Figure 3), whereas other cardiomyocytespecific genes such as ANP, Gata4 and Troponin-I were only detected in induced cells. Induced cells treated for 21 days without 5-azacytidine also expressed ANP, Gata4 and Connexin 43. We could detect weak expression of troponin-I (Figure 4).
Immunocytochemistry examination clearly detected thelocalizations of cardiac alpha-actinin and cardiac troponin I on cardiomyocytes like cells (Figure 5A and B).
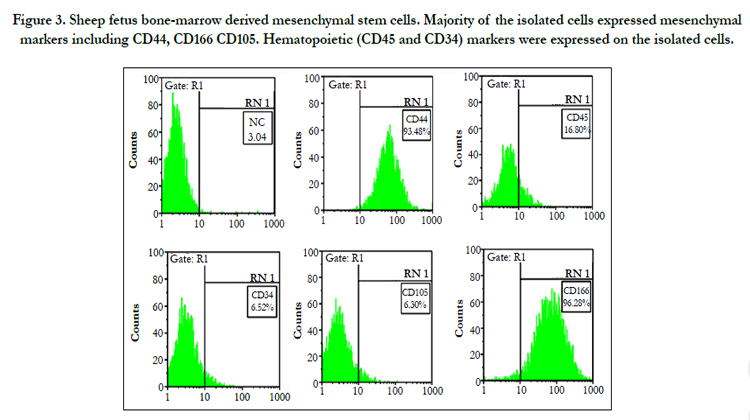
Figure 3. Sheep fetus bone-marrow derived mesenchymal stem cells. Majority of the isolated cells expressed mesenchymal markers including CD44, CD166 CD105. Hematopoietic (CD45 and CD34) markers were expressed on the isolated cells.
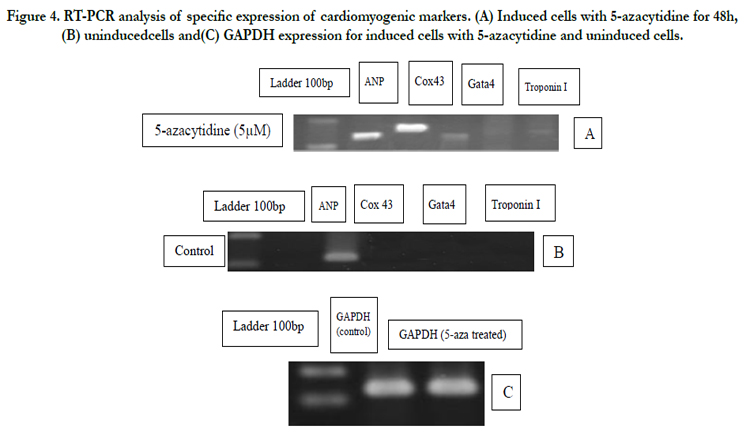
Figure 4. RT-PCR analysis of specific expression of cardiomyogenic markers. (A) Induced cells with 5-azacytidine for 48h, (B) uninducedcells and(C) GAPDH expression for induced cells with 5-azacytidine and uninduced cells.
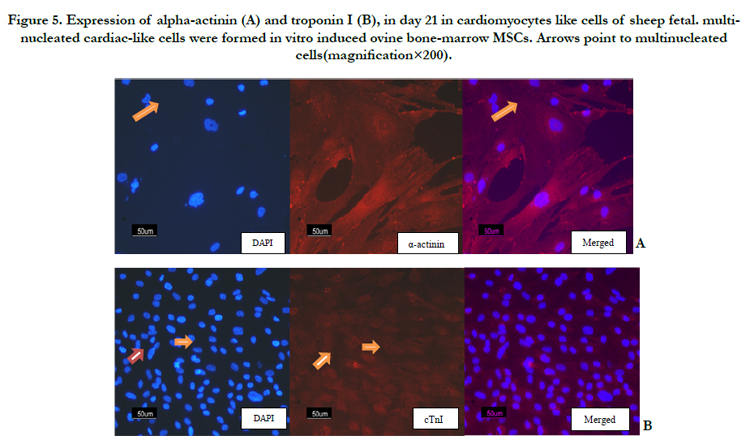
Figure 5. Expression of alpha-actinin (A) and troponin I (B), in day 21 in cardiomyocytes like cells of sheep fetal. multinucleated cardiac-like cells were formed in vitro induced ovine bone-marrow MSCs. Arrows point to multinucleated cells(magnification×200).
Discussion
MSCs isolated from animals such as mice, pigs, sheep, goats, horses, and dogs are used in preclinical trials and in the veterinarian clinic for the treatment of conditions such as osteoarthritis in dogs and musculoskeletal conditions in horses.
One of the difficulties for characterization of MSCs isolated from ruminants is lack of accessibility of specific monoclonal antibodies [12]. Godoy et al., found that sheep MSCs isolated from bone marrow, synovial membrane and adipose tissue are positive for CD44 and MHC class I [13]. In our study, flow cytometry analysis for surface markers showed that bone marrow sheep MSCs were positive for CD44 and CD166 but the expression of CD105 was weak. Despite the widespread use of MSCs from non-humans, there are no established minimal criteria for the identification of MSCs in non-humans. While all MSCs isolated from non-human display tri-lineage differentiation and plastic adherence, not all express the same panel of surface antigens that has been described for human MSCs. The expression of CD29 and CD44 in most non-human MSCs observed but the expression of some of the CD markers such as CD90, and CD105 varies depending on species and strain [14].
Most of studies have examined differentiation of MSCs into cardiomyocytes that have been treated with specific growth factors [15] and histone modifiers [16]. 5-azacytidine as a DNA methyltransferase inhibitor can affect DNA and histone methylation and causes DNA hypomethylation. Epigenetics drugs, such as 5-azacytidine can manage gene silencing activity [17], the activity of this drug may cause upregulating the differentiation-promoting factors of stem cells. The accurate function of the 5-azacytidine on cardiomyocytes differentiation from MSCs remains controversial.
The results of previous studies showed that after treatment with 5-azacytidine, differentiation rates of adult and fetal MSCs from different species is varies. The study of Zhang et al., showed that supplementation of 10 μmol/l of 5-azacytidine in culture medium of human fetal MSCs for 21 days decrease proliferation of but increases differentiation rate of human fetal MSCs into cardiomyocyte- like cells compared with 24-48h induction [18]. Antonitsisa et al., studied the addition of 5-azacytidine to the culture medium for 24h could be differentiated adult mesenchymal stem cells to cardiomyocytes [8]. Liu et al., found that, presence of 5-azacytidine in the culture medium of rat MSCs, neither the spontaneously beating cells nor the formation of myotubes were found in the primary and first passaged [1]. The presence of 5-azacytidine in the culture medium of adipose-derived stem cells (ASCs) toward cardiomyocytes differentiation was not enough [19]. Supokawej et al., proved that the addition of 15 μM 5-azacytidine could promote the expression of skeletal and cardiac muscle-specific genes to human bone marrow MSCs in culture medium [20].
In our study, the addition of 5-azacytidine to the culture medium for 48h days and then 21 days culture without this drug was caused expression of Gata4 a myocardial transcript factor in the early stage of cardiac differentiation, connexin 43 and ANP can expression in secondary stage of cardiac differentiation but we could see weak expression of troponin I. We saw the expression connexin 43 in the uninduced cells, on the basis of literature connexin 43 during embryogenesis expressed and was maintained in many different cell types [21]. Connexin 43 for multipotency and cellular migration of the skin-derived stem cell was required [21]. Although the mechanisms of 5-azacytidine toward the cardiomyocytes differentiation are still unclear, it has been proposed that this epigenetic drug may activate some silent genes by inhibiting DNA methylation [22] or induced in some genes of signaling pathways related to differentiation [23]. We think that the time of cardiac differentiation (21 days) for expression of cardiac-specific genes when 5-azacytidine for 48h added in the culture medium of sheep fetal bone marrow mesenchymal stem cells was enough. Further study is needed to elucidate the best conditions to differentiate the sheep fetal MSCs into mature cardiogenic cells.
Conclusion
In conclusion, taken together we indicated that 5-azacytidine at 5μM could enhance sheep fetal bone marrow MSC differentiation into like cardiomyocytes.
Acknowledgement
We thank Mr. Jedi the headman of Rock slaughterhouse for generous support by providing the samples.
References
- Liu Y, Song J, Liu W, Wan Y, Chen X, et al., (2003) Growth and differentiation of rat bone marrow stromal cells: does 5- azacytidine trigger their cardiomyogenic differentiation? Cardiovasc Res 58(2): 460–8.
- Klug MG, Soonpaa MH, Koh GY, Field LJ (1996) Genetically selected cardiomyocytes from differentiating embryonic stem cells form stable intracardiac grafts. J Clin Invest 98(1): 216–224.
- Wang JS, Shum-Tim D, Galipeau J, Chedrawy E, Eliopoulos N, et al., (2000) Marrow stromal cells for cellular cardiomyoplasty: feasibility and potential clinical advantages. J ThoracCardiovascSurg 120(5): 999–1005.
- Kamihata H, Matsubara H, Nishiue T, Fujiyama S, Tsutsumi Y, et al., (2001) Implantation of bone marrow mononuclear cells into ischemic myocardium enhances collateral perfusion and regional function via side supply of angioblasts, angiogenic ligands, and cytokines. Circulation 104(9):1046–1052.
- Ding S, Schultz PG (2004) A role of chemistry in stem cell biology. Nat Biotechnol 22(7): 833–840.
- Dimarakis I, Levicar N, Nihoyannopoulos P, Gordon MY, Habib NA (2006) In vitro stem cell differentiation into cardiomyocytes. Part 2: chemicals, extracellular matrix, physical stimuli and coculture assays. J Cardiothoracicrenal Res 1(2): 115–121.
- Makino S, Fukuda K, Miyoshi S, Konishi F, Kodama H, et al., (1999) Cardiomyocytes can be generated from marrow stromal cells in vitro. J Clin Invest 103(5): 697–705.
- Antonitsisa P, Ioannidou-Papagiannakib E, Kaidoglouc A, Papakonstantinoua C (2007) In vitro cardiomyogenic differentiation of adult human bone marrow mesenchymal stem cells. The role of 5-azacytidine.InteractCardiovascThoracSurg 6(5): 593–597.
- Cao F, Nic L, Meng L, Zhao L, Wang D, et al., (2004) Cardiomyocyte-like differentiation of human bone marrow mesenchymal stem cells after exposure to 5-azacytidine in vitro. J GeriatrCardiol 1(2): 101-107.
- Fukuda K (2001) Development of regenerative cardiomyocytes from mesenchymal stem cells for cardiovascular tissue engineering. Artif Organs 25(3): 187–193.
- De KA, Malakar D, Akshey YS, Jena MK , Dutta R (2011) Isolation and Characterization of Embryonic Stem Cell-Like Cells From in vitro Produced Goat (Capra hircus) Embryos. AnimBiotechnol 22(4): 181–196.
- de MattosCarvalho A, Alves AL, Golim MA, Moroz A, Hussni CA, et al., (2009) Isolation and immunophenotypic characterization of mesenchymal stem cells derived from equine species adipose tissue. Vet ImmunolImmunop 132(2): 303–306.
- Godoy RF, Alves ALG, Gibson AJ, Lima EMM, Goodship AE (2014) Do progenitor cells from different tissue have the same phenotype? Res Vet Sci 96(3), 454-459.
- Anderson P, Carrillo-Gálvez AB, García-Pérez A, Cobo M, Martín F (2013) CD105 (Endoglin)-negative murine mesenchymal stromal cells define a new multipotent subpopulation with distinct differentiation and immunomodulatory capacities. PLOS ONE 8(10): e76979.
- Bhang SH, Gwak SJ, Lee TJ, Kim SS, Park HH, et al., (2010) Cyclic mechanical strain promotes transforming-growth-factor-beta1-mediated cardiomyogenic marker expression in bone-marrow derived mesenchymal stem cells in vitro. BiotechnolApplBiochem 55(4): 191-197.
- Chang CP, Bruneau BG (2012) Epigenetics and cardiovascular development. Annu Rev Physiol 74: 41-68.
- Laurent L, Wong E, Li G, Huynh T, Tsirigos A, et al., (2010) Dynamic changes in the human methylene during differentiation. Genome Res 20(3): 320-331.
- Zhang Y, Chu Y, Shen W, Dou Z (2009) Effect of 5-azacytidine induction duration on differentiation of human first-trimester fetal mesenchymal stem cells towards cardiomyocyte-like cells. Interact Cardio VascThoracSurg 9(6): 943–946.
- Wan Safwani WKZ, Makpol S, Sathapan S, Chua KH (2012) 15-Azacytidine is insufficient for cardiogenesis in human adipose-derived stem cells. J Negat Results Biomed 11(1): 3.
- Supokawej A, Kheolamai P, Nartprayut K, U-pratya Y, Manochantr S, et al., (2013) Cardiogenic and myogenic gene expression in mesenchymal stem cells after 5-azacytidine treatment. Turk J Hematol 30(2): 115-121.
- Dyce PW, Li D, Barr KJ, Kidder GM (2014) Connexin43 is required for the maintenance of multipotency in skin-derived stem cells. Stem Cells Dev 23(14): 1636-1646.
- Chiu CP, Blau HM (1985) 5-Azacytidine permits gene activation in a previously noninducible cell type. Cell 40(2): 417–424.
- Qian Q, Qian H, Zhang X, Zhu W, Yan Y, et al., (2012) 5-Azacytidine induces cardiac differentiation of human umbilical cord-derived mesenchymal stem cells by activating extracellular regulated kinase. Stem cells dev 21(1): 67-75.





