Effects of 2-methoxyestradiol administration in mouse models purported to show signs of preeclampsia and fetal growth restriction
Poudel R1, Andersson IJ1, Stanley JL1,6,7, Rueda-Clausen CF1, Xia Xu2, Veenstra TD3, Qi H3, Sibley CP4, Davidge ST5, Baker PN6,7
1 Department of Medicine, University of Alberta, Edmonton, Alberta, Canada.
2 Frederick National Laboratory for Cancer Research, Frederick, MD, USA.
3 Department of Obstetrics and Gynecology, Chongqing Medical University, Chongqing, China.
4 Maternal and Fetal Health Research Centre, University of Manchester and Central Manchester University Hospitals NHS Foundation Trust, Manches
ter Academic Health Sciences Centre, UK.
5 Women and Children’s Health Research Institute, Edmonton, Alberta, Canada.
6 Liggins Institute, University of Auckland, Auckland, New Zealand.
7 Gravida: National Centre for Growth and Development, Auckland, New Zealand.
*Corresponding Author
Joanna L Stanley.
Liggins Institute, University of Auckland,
Auckland, 1023, New Zealand.
Tel: (64) 9 923 4616
Fax: (64) 9 373 7039
E-mail: j.stanley@auckland.ac.nz
Article Type: Case Report
Recieved: November 04, 2014; Accepted: November 21, 2014; Published: December 10, 2014.
Citation: Stanley JL et al., (2014) Effects of 2-methoxyestradiol administration in mouse models purported to show signs of preeclampsia andfetal growth restriction. Int J Reprod Fertil Sex Health, 1(5) 28-36. doi: dx.doi.org/10.19070/2377-1887-140006
Copyright: Stanley JL© 2014. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
Plasma concentration of 2-methoxyestradiol (2-ME) and placental expression of catechol-O-methyltransferase(COMT) are elevated in normal pregnancy, but reduced in women with preeclampsia (PE). Women with PE and fetal growth restriction (FGR)exhibit decreased utero-placental blood flow. Catechol-O-methyltransferase knockout mice (COMT-/-) and female CBA/J mice mated with male DBA/2 mice (CBA/J × DBA/2)show many signs of PE during pregnancy and deliver growth-restricted pups. 2-ME is known to induce significant vasorelaxation of uterine arteries from pregnant mice. We hypothesized that 2-ME administration during pregnancy will lead to an increase in uterine artery blood flow velocity in COMT-/-and CBA/J × DBA/2 mice and therefore ameliorate PE-like signs and rescue fetal growth in these animal models.
Pregnant COMT-/- and their controls (C57BL/6),and CBA/J × DBA/2 and their controls (CBA/J × BALB/c),were injected subcutaneously daily with 10 ng of 2-ME or vehicle (olive oil) from gestational day (GD) 12.5 to 17.5.
There was no significant effect of genotype/strain or treatment on maternal blood pressure, uterine artery blood flow velocity or fetal growth. A perturbed phenotype only occurred in the COMT-/- mice in regard to umbilical artery flo w velocity, proteinuria and 2-ME levels. Administration of 2-ME led to normalization of umbilical artery blood flow velocity and proteinuria in COMT-/- mice.
The utility of 2-ME in cases of PE and FGR needs to be further explored in PE models that exhibit more severe disease phenotypes.
2.Introduction
3.Materials and Methods
3.1.Animals and treatment
3.2.Blood pressure measurement
3.3.Assessment of proteinuria
3.4.Uterine and umbilical blood flow velocities
3.5.Measurement of serum 2-ME concentration
3.6.Fetal and placental measurements
3.7.Assessment of uterine artery function ex vivo
3.8.Statistical analysis
4.Results
4.1.Effect of 2-ME on blood pressure and proteinuria in COMT-/- and CBA/J × DBA/2 mice
4.2.Effect of 2-ME on uterine and umbilical artery blood flow in COMT-/-and CBA/J × DBA/2 mice
4.3.Fetal and placental measurements
4.4.Effect on ex vivo uterine artery function in COMT-/- mice
4.5.2-ME measurement in the serum
5.Discussion
6.Acknowledgements
7.References
Keywords
Pre-eclampsia; Fetal growth restriction; Mouse; 2-methoxyestradiol.
Introduction
Preeclampsia (PE) and fetal growth restriction (FGR) complicate over 10% of human pregnancies and contribute significantly to fetal and maternal morbidity and mortality [1]. PE is a condition in pregnancy characterized by high blood pressure and proteinuria after the twentieth week of pregnancy [2]. FGR is defined as the inability of a fetus to grow to its genetically determined potential size [3]. Even though PE and FGR have been known for centuries,there is still no effective treatment available, and in severe cases of PE delivery of the baby (irrespective of gestational age) remains the only cure [4].
The cause of PE and FGR is not well understood, but both conditions are known to be associated with increased uterine artery resistance [5]. In normal pregnancies, adequate trophoblast invasion of spiral arteries leads to dilation of utero-placental arteries [6]. This vascular change enables the increase of blood flow to the placental bed and, therefore, fulfills the increased demand for oxygen and nutrients from the developing fetus [7]. PE and FGR have been associated with decreased trophoblast invasion of the maternal spiral arteries [5,8,9], leading to increased resistance. As a result, uterine artery blood flow is impaired, and placental perfusion is reduced leading to ischemia. Thus, treatments directed at improving uterine artery blood flow have potential as therapies to ameliorate PE and FGR.
Although a range of murine models are available to study PE and FGR, none exhibit every facet of PE or FGR. Consequently, the two models used in this study were chosen as they exhibited evidence of both impaired uteroplacental vascular development and perfusion. The models utilized were a well-studied model of immunologically mediated pregnancy loss, female CBA/J mice mated with male DBA/2 mice (CBA/J × DBA/2), and the catechol- O-methyltransferase-deficient mouse (COMT-/-).
Decreased trophoblast number and abnormal differentiation, reduced placental perfusion and FGR have previously been observed in CBA/J x DBA/2 mice [10,11]. In addition, these mice exhibit proteinuria and renal glomerulo-endotheliosis and, therefore, appear to bean appropriate model of human PE [12]. Pregnant COMT-/- mice do not produce 2- methoxyestradiol (2-ME) and exhibit many features of PE, including raised maternal blood pressure, proteinuria, placental abnormalities, endothelial cell activation and premature delivery [13]. Kanasaki et al., (2008) reported that administration of 2-ME reversed all the features of PE in these mice [13]. Recently, we demonstrated that pups from COMT-/- mice exhibited FGR [14]. In addition, these mice exhibited abnormal umbilical artery waveforms that were characterized by reduced umbilical artery blood flow velocity and reverse or absent end-diastolic umbilical flow. These characteristics suggest that there was increased resistance in the placenta in this model. Further more, in ex vivo studies of vessel function, our laboratory has shown that uterine arteries from pregnant COMT-/- mice exhibited increased constriction in response to phenylephrine, when compared to arteries from control mice [14].
2-ME is an endogenous metabolite of estrogen that is synthesized by COMT. Plasma levels of COMT and 2-ME are typically elevated in the third trimester of normal pregnancy, but reduced in women with severe PE [13]. A recent study in Norwegian women reported that a low COMT activity haplotype was associated with recurrent PE [15]. Therefore, an alteration in the COMT gene may lead to reduced COMT enzyme activity and account for the low levels of 2-ME observed in women with PE [15]. In fact, placental COMT levels are reduced in the placentas women with severe PE [16]. In addition, reduced COMT activity has also been linked to FGR [17] which is often associated with pregnancies complicated by PE.
Preliminary studies in our laboratory have shown that 2-ME is able to induce significant vasodilation of uterine arteries (ex vivo) from normal pregnant mice and myometrial arteries from pregnant women [18]. Uterine artery endothelial cells are known to express the COMT enzyme which is required for 2-ME synthesis [19]. Further more, recent evidence suggests that 2-ME might be necessary for cytotrophoblast invasion of the maternal decidua, and, therefore, may contribute to the prevention of PE and FGR by promoting normal placental vascular formation [20]. These studies suggest that 2-ME may be able to improve uterine artery vasodilation and, therefore, uterine artery resistance in women with PE and FGR.
We hypothesized that 2-ME administration during pregnancy will lead to an increase in uterine artery blood flow velocity in murine models of PE and FGR(COMT-/- and CBA/J X DBA/2 mice) and therefore ameliorate PE-like signs and rescue fetal growth in these mice.
Materials and Methods
The University of Alberta Health Sciences Animal Policy and Welfare Committee approved all experimental procedures in this study in accordance with the guidelines of Canadian Council on Animal Care and conformed to the Guide for the Care and Use of Laboratory Animals [21].
All animals used in this study were housed in polypropylene cages in a temperature and humidity controlled environment with 12h-12h light-dark cycle. In addition, animals were given access to adlibitum standard laboratory chow and water. In this study, female COMT-/-mice (Courtesy of Professor J Gogos, University of Columbia) and their respective controls (C57BL/6J mice)(Jackson Laboratories; Bar Harbor, ME) of 2-3 months of age were nightly mated with strain-matched males. CBA/J females were mated with DBA/2 males and as controls CBA/J were mated with BALB/c males (Jackson Laboratories; Bar Harbor, ME). The day of vaginal plug detection was designated as 0.5 day of pregnancy. From gestational day (GD) 12.5 to GD 18.5, pregnant dams were injected subcutaneously every day with 10 ng (100 μL) of 2-ME or vehicle (olive oil 100 μL). The timing of the dose was based on previous studies, in which we used other interventions in mouse models using an identical time frame and were able to rescue fetal growth [14,22]. The selected dose of 2-ME was based on previous studies [13,20]. The dose administered in the current study increased the serum concentration of 2-ME in COMT-/- mice to serum concentrations similar to those observed in the control wild-type pregnant mice [13].
Systolic blood pressure was measured using non-invasive tail-cuff method (IITC Life Science, Woodland Hills, CA, USA) on GD 17.5.
Urine samples were collected on GD 18.5 of pregnancy. Samples were then stored at -80°C until assayed for urinary albumin using an enzyme-linked immunosorbent assay (ELISA) (AssayPro, St Charles, MO, USA) and urinary creatinine by colorimetric kit assay (Cayman Chemical Company, Ann Arbor, MI, USA). From these measurements the albumin to creatinine ratio was calculated as μg albumin/mg creatinine.
Uterine and umbilical artery and vein blood flow velocity were assessed in vivo on GD 17.5. Blood flow velocities were measured using previously established protocols [14,23]. Briefly, mice were anesthetized with isofluorane (3%) in air and placed in a supine position on a temperature-controlled heating pad. Heating was adjusted to maintain rectal temperature between 36°Cand 38°C. Then the anesthetic concentration was reduced (to ~0.5% to 1.5%) in order to maintain the constant maternal heart rate of 550 ± 50 bpm and a respiratory rate of 150 ± 20 cpm. Abdominal hair was removed by the application of a chemical hair remover. Prewarmed gel was applied to the shaved abdomenas an ultrasoundcoupling medium before mice were imaged transcutaneously using a high-resolution ultrasound biomicroscope (model Vevo 770 for COMT-/- and C57BL6/J mice and model Vevo 2100 for CBA/J × DBA/2 and CBA/J X BALB/c mice;VisualSonics®, Toronto, ON, Canada). The transducer used for Vevo 770 was 30 MHz operating at 100 frames/s. The Vevo 2100 system was also equipped with a 30 MHz transducer but it was operating at 300 frames/s. The angle between the Doppler beam and the vessel was <30 degrees. Doppler waveforms were then obtained from both left and right uterine artery near the uterocervical junction close to the iliac artery. Umbilical artery and vein waveforms were obtained from at least two fetuses near the placental surface. Peak systolic velocity (PSV) and end diastolic velocity (EDV) were measured from at least three consecutive cardiac cycles that were not affected by motion caused by maternal breathing and the results were averaged. The resistance index (RI) = ((PSV-EDV)/ PSV) was calculated.
On GD18.5, mice were subjected to terminal anesthesia using isofluorane and blood was withdrawn by heart puncture. Blood was then allowed to clot for ~1h in ice. Serum was then extracted following centrifugation for 10 minutes at 5000 rpm and stored in -80°C. The serum 2-ME concentration at GD 18.5 was measured in the COMT -/-, COMT-/- mice supplemented with 2-ME and C57BL6/J mice by liquid chromatography-mass spectroscopy as previously described [24].
Dams were euthanized on GD 18.5 and the entire uterus was removed and immediately placed in freshly prepared cold physiological saline solution (PSS). Pups and placentas were then dissected out and number of live fetuses and resorptions were recorded. The fetuses and placentas were blotted dry and weighed, and the crown to rump length and abdominal circumference were recorded. Placentas were further dried at 37°C and the dry placental weight was recorded. Uterine arteries from the right horn were carefully dissected in cold PSS and surrounding adipose and connective tissue was removed.
Uterine arteries were carefully cut into four small segments and were mounted into a wire myograph (610M, Danish Myo Technology, Aarhus, Denmark) as described elsewhere [14,25]. Briefly, the vessels were secured in 25μm tungsten wire and then warmed to 37°C (bathed in 6 mLof PSS) for 20 minutes before they were normalized to 0.9 of L13.3kPa using standard normalization procedure. Vessel function was then assessed by measuring contraction and relaxation response curves in response to phenylephrine (Phe) and methacholine (Mch), respectively, as described previously [14]. Briefly, a cumulative dose response curve (0.1 nmol/L - 10μmol/L) was performed for Phe. The Phe concentration required to produce 80% constriction or EC80was then calculated and added to the bath and the endothelium-dependent relaxation response was tested by subsequent vasodilation curve to MCh (0.1 nmol/L - 10μmol/L).
All data is presented as mean ± standard error of the mean (SEM) and compared with two-way analysis of variance that included genotype and resveratrol treatment as independent variables. Significance was determined using the Bonferroni post-hoc test and statistical significance was defined as P<0.05. Graphpad Prism (La Jolla, CA) was used for statistical analysis. For wire myography experiments, sigmoidal dose response curve fittings were performed and EC50 concentration of the drug was calculated using Graphpad Prism 5.0.
Results
Systolic blood pressure (SBP) and heart rate (HR) on GD 17.5 were not different between the C57BL/6J and COMT-/- mice(Table 1). In addition, there was no difference in SBP and HR between CBA/J × DBA/2 and their respective controls CBA/J X BALB/c (Table 1).
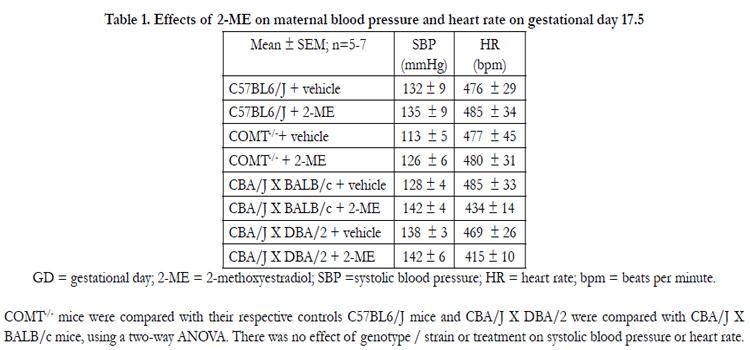

Table 1. Effects of 2-ME on maternal blood pressure and heart rate on gestational day 17.5
GD = gestational day; 2-ME = 2-methoxyestradiol; SBP =systolic blood pressure; HR = heart rate; bpm = beats per minute.
COMT-/- mice were compared with their respective controls C57BL6/J mice and CBA/J X DBA/2 were compared with CBA/J X
BALB/c mice, using a two-way ANOVA. There was no effect of genotype / strain or treatment on systolic blood pressure or heart rate.
There was a significant effect of genotype and treatment on albumin:creatinine ratio. In mice treated with vehicle, the albumin:creatinine ratio was significantly elevated in the COMT-/- mice compared to C57BL/6J (control mice) at GD 18.5 (Figure 1A; P<0.05). Following treatment with 2-ME, the albumin:creatinine ratio was reduced in the COMT-/- mice and was similar to those recorded in the control animals treated with vehicle (Figure 1A; P<0.05, COMT-/- vehicle-treated mice vs COMT-/- 2 ME-treated mice). However, proteinuria was not significantly different between CBA/J × DBA/2 and CBA/J X BALB/c mice and 2-ME treatment did not have any effect on this measure in these groups of mice (Figure 1B).
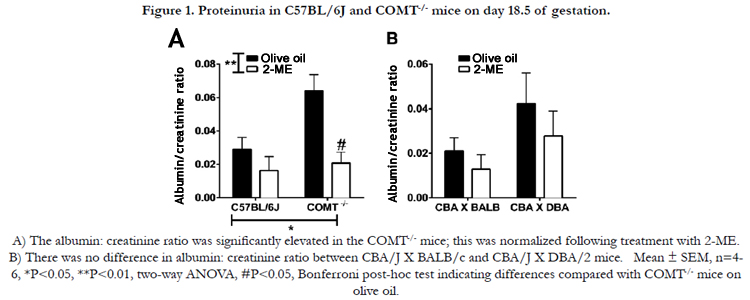
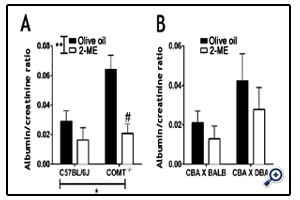
Figure 1. Proteinuria in C57BL/6J and COMT-/- mice on day 18.5 of gestation.
A) The albumin: creatinine ratio was significantly elevated in the COMT-/- mice; this was normalized following treatment with 2-ME.
B) There was no difference in albumin: creatinine ratio between CBA/J X BALB/c and CBA/J X DBA/2 mice. Mean ± SEM, n=4-
6, *P<0.05, **P < 0.01, two-way ANOVA, #P < 0.05, Bonferroni post-hoc test indicating differences compared with COMT-/- mice onolive oil.
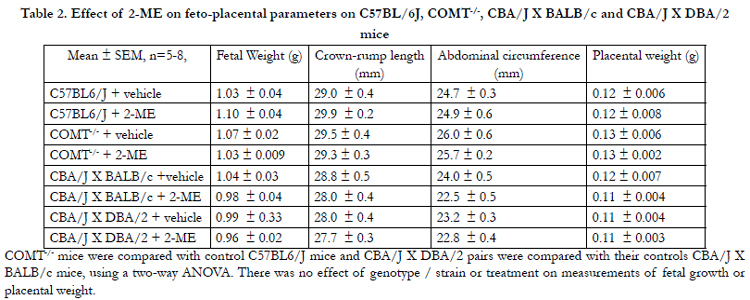
Table 2. Effect of 2-ME on feto-placental parameters on C57BL/6J, COMT-/-, CBA/J X BALB/c and CBA/J X DBA/2 mice
COMT-/- mice were compared with control C57BL6/J mice and CBA/J X DBA/2 pairs were compared with their controls CBA/J X BALB/c mice, using a two-way ANOVA. There was no effect of genotype / strain or treatment on measurements of fetal growth or placental weight.
The hemodynamic and waveform parameters, as measured at GD 17.5, are listed in Table 3 and 4. There were no significant differences in uterine artery and umbilical vein blood flow velocities in C57BL/6J and COMT-/- mice at GD 17.5 (Table 3). Umbilical artery maximum velocities were also not significantly different between C57BL/6J and COMT-/- mice (Figure 2A). However, a significant effect of genotype and treatment on umbilical artery end diastolic velocity and resistance index was observed on GD 17.5 in COMT-/- mice (P<0.05, Figure 2B). The minimum umbilical artery velocity was significantly reduced (P<0.05, Figure 2B) and the resistance index was increased in the COMT-/- mice (Figure 2C). Importantly, both of these measures were normalized following treatment with 2-ME in the COMT-/- mice (Figure 2B and 2C).
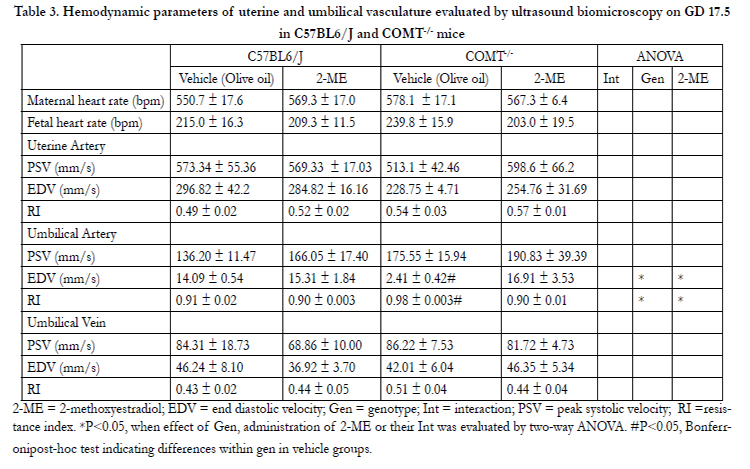
Table 3. Hemodynamic parameters of uterine and umbilical vasculature evaluated by ultrasound biomicroscopy on GD 17.5 in C57BL6/J and COMT-/- mice
2-ME = 2-methoxyestradiol; EDV = end diastolic velocity; Gen = genotype; Int = interaction; PSV = peak systolic velocity; RI =resistance index. *P<0.05, when effect of Gen, administration of 2-ME or their Int was evaluated by two-way ANOVA. #P<0.05, Bonferronipost-hoc test indicating differences within gen in vehicle groups.
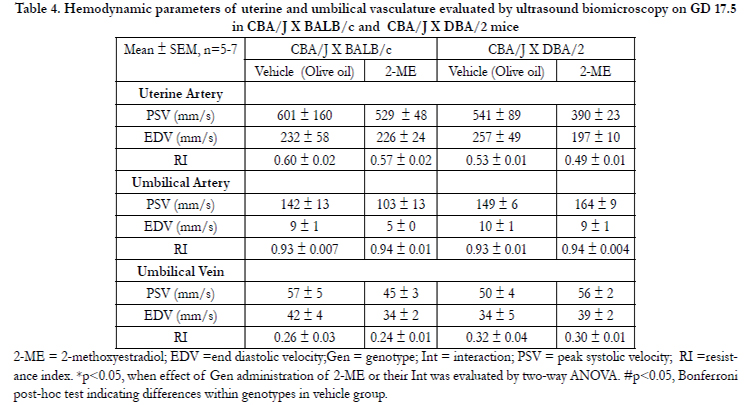
Table 4. Hemodynamic parameters of uterine and umbilical vasculature evaluated by ultrasound biomicroscopy on GD 17.5 in CBA/J X BALB/c and CBA/J X DBA/2 mice
2-ME = 2-methoxyestradiol; EDV =end diastolic velocity;Gen = genotype; Int = interaction; PSV = peak systolic velocity; RI =resistance index. *p<0.05, when effect of Gen administration of 2-ME or their Int was evaluated by two-way ANOVA. #p<0.05, Bonferroni post-hoc test indicating differences within genotypes in vehicle group.
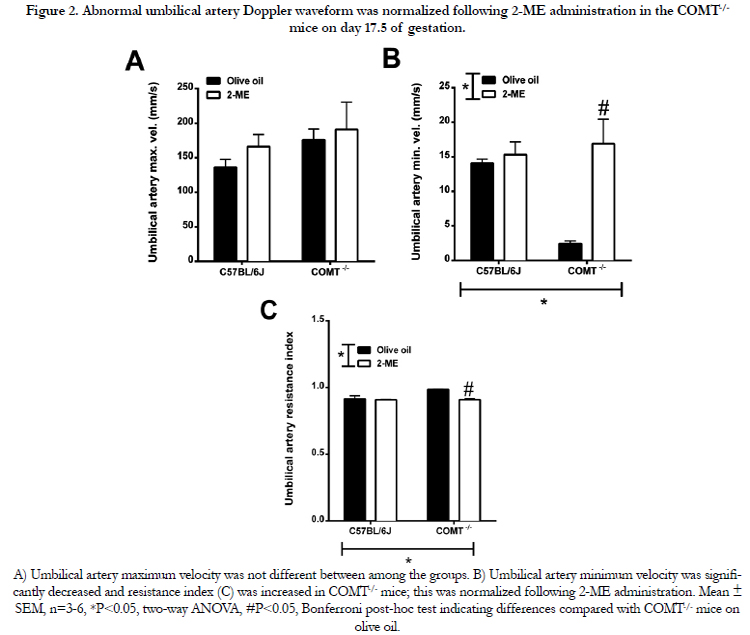
Figure 2. Abnormal umbilical artery Doppler waveform was normalized following 2-ME administration in the COMT-/- mice on day 17.5 of gestation.
A) Umbilical artery maximum velocity was not different between among the groups. B) Umbilical artery minimum velocity was significantly decreased and resistance index (C) was increased in COMT-/- mice; this was normalized following 2-ME administration. Mean ± SEM, n=3-6, *P<0.05, two-way ANOVA, #P<0.05, Bonferroni post-hoc test indicating differences compared with COMT -/- mice on olive oil.
There was no significant difference in pup weight, crown-rump length, or abdominal circumference between C57BL/6J and COMT-/- mice (Table 2). These parameters were also not significantly different between CBA/J × DBA/2 and CBA/J X BALB/c mice (Table 2). In addition, 2-ME did not have any effect on any of these measurements in either of the mice models (Table 2). Placental weights were not significantly different between animals in any of the groups. Similarly, the pup weight:placental weight ratio was not significantly different between the two murine model and their respective controls; 2-ME did not have any effect on this measure (Table 2). There was no evidence of gross abnormalities in fetuses from 2-ME-treated dams.
There was no effect of genotype or 2-ME treatment on Pheinduced constriction in COMT-/-(Figure 3A) and CBA/J × DBA/2mice (Figure 4A). Similarly, there was no effect of genotype or 2-ME treatment on sensitivity to Phe either in COMT-/- (Figure 3B) or in CBA/J × DBA/2mice (Figure 4B). Additionally, MCh-induced relaxation or MCh EC50 concentrations were not significantly different between C57BL/6J and COMT-/- mice (Figure 3C) and CBA/J × DBA/2 and CBA/J X BALB/c mice (Figure 4C). Supplementation with 2-ME did not have any effect on these measuresin COMT-/- mice (Figure 3D) and CBA/J × DBA/2 and CBA/J X BALB/c mice (Figure 4D). No significant difference in uterine artery relaxation was observed among the mice models and their respective controls in response to the NO donor SNP (data not shown).
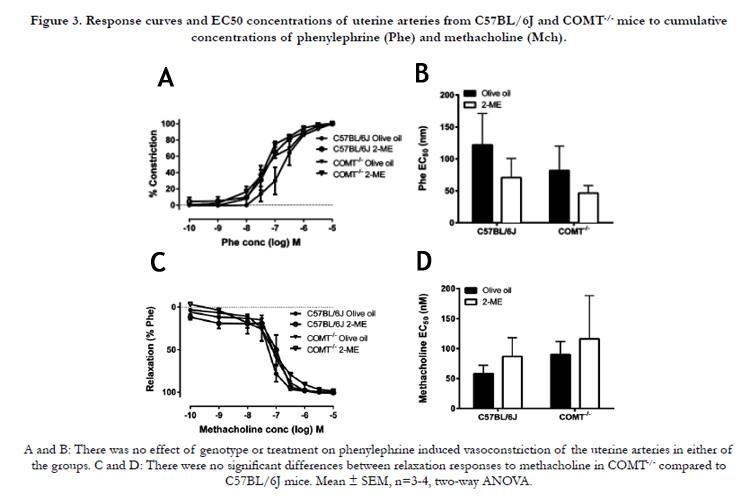
Figure 3. Response curves and EC50 concentrations of uterine arteries from C57BL/6J and COMT-/- mice to cumulative concentrations of phenylephrine (Phe) and methacholine (Mch).
A and B: There was no effect of genotype or treatment on phenylephrine induced vasoconstriction of the uterine arteries in either of the groups. C and D: There were no significant differences between relaxation responses to methacholine in COMT-/- compared to C57BL/6J mice. Mean ± SEM, n=3-4, two-way ANOVA.
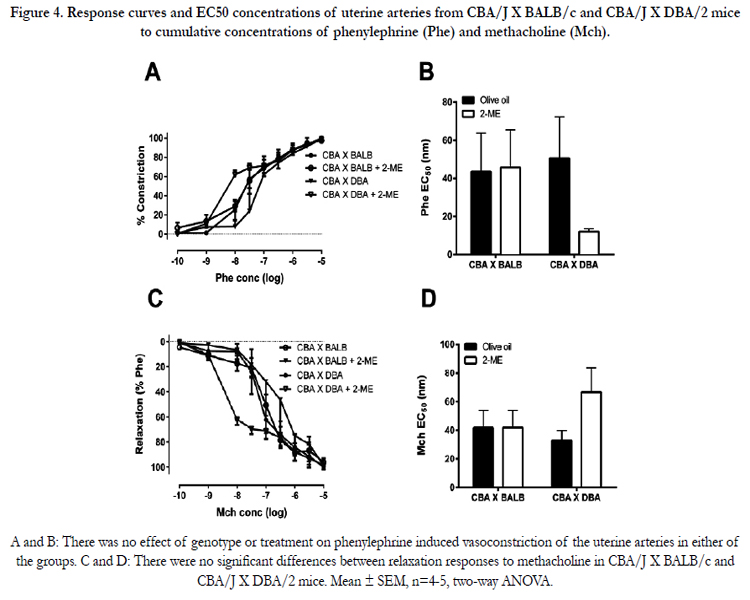
Figure 4. Response curves and EC50 concentrations of uterine arteries from CBA/J X BALB/c and CBA/J X DBA/2 mice to cumulative concentrations of phenylephrine (Phe) and methacholine (Mch).
A and B: There was no effect of genotype or treatment on phenylephrine induced vasoconstriction of the uterine arteries in either of the groups. C and D: There were no significant differences between relaxation responses to methacholine in CBA/J X BALB/c and CBA/J X DBA/2 mice. Mean ± SEM, n=4-5, two-way ANOVA.
Serum concentration of 2-ME was significantly reduced in COMT-/- mice compared with C57BL/6J control mice (0.033 ± 0.007 vs. 0.085 ± 0.021 pg/ml; p<0.05). Administration of 2-ME increased circulating concentration of this molecule in COMT-/- mice to a level that was not significantly different to the level of 2-ME found in control mice (0.130 ±0.019 pg/ml; p>0.05).
Discussion
It is known that 2-ME induces significant vasodilation of uterine arteries from normal pregnant mice [18] and of aortic segments from male rats ex vivo [26]. In addition, 2-ME is known to ameliorate high blood pressure and proteinuria in COMT-/- mice during pregnancy [13]. We hypothesized that treatment of COMT-/- and CBA/J × DBA/2mice with 2-ME during pregnancy would improve uterine artery blood flow velocity and, therefore, ameliorate signs of PE and rescue fetal growth. In the current study, we observed that 2-ME administration during pregnancy did not increase uterine artery blood flow velocity in the COMT-/- and CBA/J × DBA/2 mice. However, following 2-ME administration, the increased proteinuria, and reduced umbilical artery blood flow velocity, found in the COMT-/- mice were normalized to that of control animals.
Since 2-ME abolishes vascular contraction in the presence, but not in the absence, of endothelium, and this effect is blocked by NOS inhibition, it suggests that 2-ME abrogates vascular contraction via endothelium-dependent NO production[26,27]. Given the capacity of 2-ME to induce vascular relaxation, its ability to increase uterine artery blood flow velocity in the COMT-/-and CBA/J × DBA/2 mice was investigated. In agreement with our previous study [14], there was no difference in uterine artery blood flow velocity in the COMT-/- compared to C57BL/6J mice and 2-ME did not have an effect on this measure. CBA/J × DBA/2 mice did not exhibit abnormal uterine artery blood flow velocity compared to their controls CBA/J X BALB/c mice. In line with blood flow results, we did not observe any differences between the normal control mice C57BL6/ and COMT-/- mice and CBA/J × DBA/2 and CBA/J X BALB/c mice in ex vivo uterine artery endothelial function studies.
In previous studies, CBA/J × DBA/2 mice have been shown to exhibit FGR [10,11]. This abnormality was not present in the current study. One of the reasons for this disparity may be due to the difference in gestational age at which the measurements were made i.e. fetal weight measured at gestational day 15, as opposed to 18.5 in this study [10,11].
COMT-/- mice have previously [14] demonstrated abnormal umbilical Doppler waveforms, including decreased minimum umbilical artery blood flow velocity and increased umbilical artery resistance. 2-ME administration led to normalization of these measures in the COMT-/- mice. Although the abnormal umbilical Doppler waveforms were associated with FGR in the COMT-/- mice in our previous study [[14], FGR was not observed in the current tudy. Similarly, Kanasaki et al., (2008) did not report FGR in the pups from COMT-/- mice [13]. It is important to note that the growth restriction observed in the pups from COMT-/- mice in our previous study was subtle [14]. In the current study, it is possible that the abnormal umbilical Doppler waveforms were present only in a subset of fetuses and, therefore, growth restriction did not appear in all fetuses. Given that 2-ME administration led to normalization of abnormal umbilical Doppler waveforms in the COMT-/- mice, its therapeutic potential in cases of FGR with abnormal umbilical Doppler waveforms is apparent. However, the mechanisms by which 2-ME normalizes umbilical Doppler waveforms in the COMT-/- mice remain to be investigated.
Placentas from COMT-/- mice exhibit an increased in the antiangiogenic factor sFlt-1[13]. This can lead to a reduction in placental angiogenesis in these mice, and consequently increase placental resistance and abnormal umbilical Doppler waveforms. One of the mechanisms by which 2-ME can lead to normalization of abnormal umbilical Doppler waveforms might be via its effect on hypoxia-inducible factor HIF-1α, a transcription factor that senses tissue oxygen tension and regulates the expression of hypoxia-induced genes. Placental resistance might be reduced by 2-ME through inhibition of HIF-1α, which is known to upregulate sFlt1 [28,29]. Another possible mechanism by which 2-ME could normalize the abnormal umbilical Doppler waveforms in the COMT-/- mice may be through restoration of the placental eNOS enzyme expression [13]. This might lead to an increase in NO production in the placental vascular bed resulting in a reduction in placental resistance. Recently, 2-ME was measured in 157 cord blood samples spanning a wide range of gestation (24 – 42 weeks) [30].It was discovered that infants born between 37 and 42 weeks had significantly increased levels of 2-ME compared to infants born at earlier gestation. Although it remains to be verified, it is plausible that 2-ME supplementation might have led to an increase in umbilical 2-ME concentrations in the COMT-/- mice. Therefore, 2-ME might elicit its vasorelaxing effects in the umbilical circulation. This could in part explain the normalization of impaired umbilical Doppler waveforms in these mice.
Kanasaki et al., (2008) first demonstrated that COMT-/- mice displayed a PE-like phenotype, with significantly elevated blood pressure and proteinuria at GD 17.5 [13]. Therapy with 2-ME led to amelioration of these signs. In agreement with this study [13], COMT-/-mice in our laboratory exhibited increased proteinuria and this was normalized to wild-type levels following 2-ME administration. In contrast to this study [13], but consistent with our previous study [14], COMT-/- mice did not display hypertension on GD 17.5. The use of different control mice in the two studies might account for the observed difference in blood pressure. Kanasaki et al., (2008) used COMT+/+ (littermates of COMT-/-) as controls in their study [13], whereas we used C57BL6/J mice, which are genetic background for the COMT-/- mice. It is important to note that COMT+/+ mice in the study by Kanasaki et al., (2008) did not display the pregnancy-induced blood pressure adaptation [13] observed in C57BL/6J mice [31] and in human pregnancy [8]. In normal pregnancy in both humans [8] and mice [31], there is a steady decrease in blood pressure up to mid-pregnancy and blood pressure returns to normal in the last weeks of pregnancy. In the COMT+/+ mice used by Kanasaki et al., (2008), blood pressure continued to decrease until the end of pregnancy [31], which might explain the blood pressure difference found between the two studies.
In summary, the current study demonstrated that 2-ME treatment in COMT-/- mice led to normalization of proteinuria and umbilical artery Doppler waveforms, suggesting that 2-ME might have therapeutic potential in cases of FGR associated with abnormal Doppler waveforms. However, in the CBA/J × DBA/2 mice, a PE-like phenotype and FGR were not found in the current study. Thus, they can likely be excluded as a model to tests drugs aimed at improving these conditions. The utility of 2-ME in cases of PE needs to be further explored in PE models that exhibit more severe disease phenotypes.
Acknowledgements
The authors would like to thank Professor J.Gogos, University of Columbia and Professor J. Waddington, Royal College of Surgeons Ireland for providing the COMT-/- mice.
References
- Steegers EA, von Dadelszen P, Duvekot JJ, Pijnenborg R (2010) Pre-eclampsia. Lancet 376: 631-644.
- ACOG Committee on Practice Bulletins-Obstetrics (2002) ACOG practice bulletin. Diagnosis and management of preeclampsia and eclampsia. Obstet Gynecol 99: 159-167.
- Figueras F, Gratacos E (2014) Update on the diagnosis and classification of fetal growth restriction and proposal of a stage-based management protocol. Fetal Diagn Ther 36: 86-98.
- Redman CW (2011) Hypertension in pregnancy: the NICE guidelines.Heart 97: 1967-1969.
- Harrington K, Cooper D, Lees C, Hecher K, Campbell S (1996) Doppler ultrasound of the uterine arteries: the importance of bilateral notching in the prediction of pre-eclampsia, placental abruption or delivery of a small-forgestational-age baby. Ultrasound Obstet Gynecol 7: 182-188.
- Hladunewich M, Karumanchi SA, Lafayette R (2007) Pathophysiology of the clinical manifestations of preeclampsia. Clin J Am Soc Nephrol 2: 543-549.
- Gagnon R (2003) Placental insufficiency and its consequences. Eur J Obstet Gynecol Reprod Biol 110: S99-S107.
- Clark SL, Cotton DB, Lee W, Bishop C, Hill T, et al. (1989) Central hemodynamic assessment of normal term pregnancy. Am J Obstet Gynecol 161:1439-1442.
- Pijnenborg R, Vercruysse L, Hanssens M (2006) The uterine spiral arteries in human pregnancy: facts and controversies. Placenta 27: 939-958.
- Girardi G, Yarilin D, Thurman JM, Holers VM, Salmon JE (2006) Complement activation induces dysregulation of angiogenic factors and causes fetal rejection and growth restriction. J Exp Med 203: 2165-2175.
- Redecha P, van Rooijen N, Torry D, Girardi G (2009) Pravastatin prevents miscarriages in mice: role of tissue factor in placental and fetal injury. Blood 113: 4101-4109.
- Ahmed A, Singh J, Khan Y, Seshan SV, Girardi G (2010) A new mouse model to explore therapies for preeclampsia. PloS One 5: e13663.
- Kanasaki K, Palmsten K, Sugimoto H, Ahmad S, Hamano Y, et al. (2008) Deficiency in catechol-O-methyltransferase and 2-methoxyoestradiol is associated with pre-eclampsia. Nature 453: 1117-1121.
- Stanley JL, Andersson IJ, Poudel R, Rueda-Clausen CF, Sibley CP, et al. (2012) Sildenafil citrate rescues fetal growth in the catechol-O-methyl transferase knockout mouse model. Hypertension 59: 1021-1028.
- Roten L, Fenstad M, Forsmo S, Johnson M, Moses E, et al. (2011) A low COMT activity haplotype is associated with recurrent preeclampsia in a Norwegian population cohort (HUNT2). Mol Hum Reprod 17: 439-446.
- Barnea E, MacLusky N, DeCherney A, Naftolin F (1988) Catechol-Omethyl transferase activity in the human term placenta. Am J Perinatol 5:121-127.
- Sata F, Yamada H, Suzuki K, Saijo Y, Yamada T, et al. (2006) Functional maternal catechol-O-methyltransferase polymorphism and fetal growth restriction.Pharmacogenet Genomics 16: 775-781.
- Stanley J, Zhang Y, Crocker I, Baker P (2010) The effect of 2-methoxyestradiol on uterine artery function. Proceedings of the Society for Gynecological Investigation Annual Meeting.
- Jobe SO, Ramadoss J, Koch JM, Jiang Y, Zheng J, et al. (2010) Estradiol- 17β and its cytochrome P450-and catechol-O-methyltransferase–derived metabolites stimulate proliferation in uterine artery endothelial cells role of estrogen receptor-β versus estrogen receptor-β Hypertension 55: 1005-1011.
- Lee SB, Wong AP, Kanasaki K, Xu Y, Shenoy VK, et al. (2010) Preeclampsia: 2-methoxyestradiol induces cytotrophoblast invasion and vascular development specifically under hypoxic conditions. Am J Pathol 176: 710-720.
- Institute of Laboratory Animal Resources (U.S.). Committee on Care and Use of Laboratory Animals. (1996) Guide for the care and use of laboratory animals. NIH publication. Bethesda, MD: U.S. Dept. of Health and Human Services, Public Health Service.
- Stanley JL, Andersson IJ, Hirt CJ, Moore L, Dilworth MR, et al. (2012) Effect of the anti-oxidant tempol on fetal growth in a mouse model of fetal growth restriction. Biol Reprod 87:25, 21-28,.
- Mu J, Adamson SL (2006) Developmental changes in hemodynamics of uterine artery, utero-and umbilicoplacental, and vitelline circulations in mouse throughout gestation. Am J Physiol Heart Circ Physiol 291: H1421-H1428.
- Xu X, Keefer LK, Ziegler RG, Veenstra TD (2007) A liquid chromatography–mass spectrometry method for the quantitative analysis of urinary endogenous estrogen metabolites. Nat Protoc 2: 1350-1355.
- Cooke CL, Davidge ST (2003) Pregnancy-induced alterations of vascular function in mouse mesenteric and uterine arteries. Biol Reprod 68: 1072-1077.
- Gui Y, Zheng XL, Zheng J, Walsh MP (2008) Inhibition of rat aortic smooth muscle contraction by 2-methoxyestradiol. Am J Physiol Heart Circ Physiol 295: H1935-H1942.
- Wenzel E, Somoza V (2005) Metabolism and bioavailability of trans‐resveratrol. Mol Nutr Food Res 49: 472-481.
- Mabjeesh NJ, Escuin D, LaVallee TM, Pribluda VS, Swartz GM, et al. (2003) 2ME2 inhibits tumor growth and angiogenesis by disrupting microtubules and dysregulating HIF. Cancer Cell 3: 363-375.
- Gerber HP, Condorelli F, Park J, Ferrara N (1997) Differential transcriptional regulation of the two vascular endothelial growth factor receptor genes. Flt-1, but not Flk-1/KDR, is up-regulated by hypoxia. J Biol Chem 272: 23659-23667.
- Barnes CM, McElrath TF, Folkman J, Hansen AR (2010) Correlation of 2-methoxyestradiol levels in cord blood and complications of prematurity.Pediatr Res 67: 545-550.
- Burke SD, Barrette VF, Gravel J, Carter AL, Hatta K, et al. (2010) Uterine NK cells, spiral artery modification and the regulation of blood pressure during mouse pregnancy. Am J Reprod Immunol 63: 472-481.





