Production and Simple Purification of Recombinant Human Granulocyte Colony-Stimulating Factor using the Inteintag in Escherichia Coli
Peymanfar P1, Roghanian R1*, Ghaedi K1, Sayed H1, Yari R2
1 Biology Department, Faculty of Science, University of Isfahan, Hezar Jaribaven, Isfahan, Iran.
2 Biology Department, Faculty of Science, Islamic Azad University, Boroujerd, Iran.
*Corresponding Author
Rasoul Roghanian,
Faculty of Science, Biology Department,
University of Isfahan, Hezar Jaribaven, Postal code:81746, Isfahan, Iran.
E-mail: rasoul_roghanian@yahoo.co.uk
Received: July 07, 2016; Accepted: July 27, 2016; Published: August 04, 2016
Citation: Peymanfar P, Roghanian R, Ghaedi K, Sayed H, Yari R (2016) Production and Simple Purification of Recombinant Human Granulocyte Colony-Stimulating Factor using the Inteintag in Escherichia Coli. Int J Med Biotechnol Genetics. 4(2), 40-46.doi: dx.doi.org/10.19070/2379-1020-160007
Copyright: Roghanian R© 2016. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
Background: Granulocyte colony-stimulating factor (G-CSF) is a cytokine which have many functions such as stimulating of hematopoiesis, proliferation and differentiation of granulocyte progenitor cells and production of bone marrow neutrophilic granulocyte colonies. Nowadays, the usage of human recombinant G-CSF (rh G-CSF) is for curing of chemotherapy-radiotherapy-induced neutropenia, and also in patients with bone marrow transplantation. The aim of this study was to produce a soluble type of G-CSF in E.coli and purification in new way. In this study, we achieved active G-CSF using the intein fusion system. Therefore, two copies of optimized Intein - GCSF genewas cloned into the pET22b vector to generate pET22b-G-CSF-intein2 (C-terminal fusion vector) and pET22b intein1-G-CSF (N-terminal fusion vector). The hG-CSF sequence with two different intein sequence from pTWIN1 vector are synthesized and then inserted into a pET22 expression vector under the control of T7 promoter and cloned in E.coli strain BL21 (DE3).The fused proteins formed misfolded proteins as inclusion bodies (IBs). The IBs were solved with urea solution and afterward proteins were saved for size-exclusion chromatography. Then , CBD-intein–GCSF was loaded onto chitin beads column equilibrated with 10mM Tris buffer, 500mM NaCl, pH 8.5, and was eluted from the column by incubation at 25°C under pH 6.5 for 16h based on intein C-terminal self-cleavage. SDS-Page, western blot, dot blot, size exclusion chromatography and in vivo assay of protein demonstrated that the bioactivity of recombinant GCSF was compared with standard GCSF.
Results: After culturing and induction of recombinant E.coli with IPTG, we obtained a good expression of the hG-CSF, as determined by SDS-PAGE and confirmed by dot blotting and western blotting. Approximately 50% of the expressed G-CSF was soluble after the IPTG induction and temperature reduction. Up to 1.2 mg of G-CSF was achieved from a 1 L culture, and nearly75% of the GCSF-Intein2 was cleared by pH shift. Intein1-GCSF was not cleaved due to the inhibition of cleavage by the N-terminal amino acid of GCSF. The biological activity assay, in vivo, showed a higher biological effect than standard reference hG-CSF.
Conclusion: The immunological and biological analyses showed that this protocol can be useful to develop bioproducts. In conclusion, the combination of different methods presented here permitted an simple and cost-effective protocol for rhG-CSF production.
2.Introduction
3.Materials and Methods
3.1.Bacterial Strains, Plasmids, and Media
3.2.Fermentation
3.3.Medium Composition and Experimental Design
3.4.Purification with On-Column Cleavage
3.5.Detection of dimers and monomers withSize Exclusion Chromatography (SEC-HPLC)
3.6.Biological Assay
4.Results and Discussion
4.1.Selection of Basal Medium
4.2.Biological potency evaluation
5.Acknowledgments & Declarations
6.References
Keywords
Granulocyte Colony Stimulating Factor; Recombinant; Recombinant Proteins; Escherichia Coli.
Introduction
E.coli cells are well characterized bacteria in terms of molecular genetics, physiology and expression systems. They are good host cells and could be engineered easily. Proteins production in E.coli has more benefits than other production ways. Strategies for improving the expression of heterologous protein in E.coli periplasmic spaces have been severly studied [1, 2]. The recombinant proteins were secreted via translocation across the cytoplasmic membrane to periplasmic space. This way has a good potential for proteins especially for pharmaceutical companies [3]. The problems of protein misfolding, insoluble aggregated proteins, and incorrect mature protein can be decreased by expression of proteins in periplasm [4, 5]. Additionally, the periplasmic space is a more oxidative and less protease activity environment compared with cytoplasm [6]. The granulocyte colony stimulating factor (G-CSF) have a regulatory role in the growth, differentiation, survival, and activation of neutrophils and their precursors in immune system [4]. Chemotherapy of cancer can prevent production of these white blood cells, leaving patients sensitive to infections. Thus ,G-CSF has been successfully used in cancer patients whose treatment needs high-dose chemotherapy [5]. Additionally, G-CSF can be used to regenerate the immune system in patients with HIV, pneumonia, diabetic foot infections, caner and febrile neutropenia [7, 8]. According to this extensive application, the recombinant human GCSF has been expressed in engineered E.coli and was approved for using in chemotherapy-induced neutropenia by the U.S Food and Drug Administration in 1991. It should be pointed out that two types of clinically available G-CSF are: a lenograstim (glycosylated form is produced in mammalian cells), and filgrastim (nonglycosylated form is produced in E.coli [9]. Both prokaryotic (E.coli) and eukaryotic (Pichia pastoris) production host systems were used to produce G-CSF and eukaryotic (Pichia pastoris and CHO cells) production host systems were used to produce G-CSF [10, 11]. Recent studies describe various protocols for cloning, expression and purification of the rhG-CSF. These protocols shows use of different chromatography columns and diverse detergents for the purification [9-11] and some of them were not applicable for purifying of recombinant G-CSF from inclusion bodies [19]. In pharmaceutical industry, production of bioproducts needs a simple and cost-effective process involving easier steps that have high yield. Here we describe the cloning of recombinant human granulocyte colony-stimulating factor gene into pET22b vector, protein expression in E.coli BL21 cells, a simple purification protocol of the active recombinant protein with inteins and chitin beads from inclusion bodies and characterization of rhGCSF by analytical methods. We believe that the combination of the different experimental strategies presented here provides an efficient protocol that may be useful in the pharmaceutical industry for rhG-CSF production.
The host cell for expression experiments was E.coli BL21 (DE3),while host for cloning of the fusion genes was E.coli DH5α. The initial cloning and protein expression was done by pET-22b (+) vector, respectively. The hG-CSF sequence and two different intein sequence from pTWIN1 vector were bought from NCBI [12, 13]. GENESCRIPT company synthesized all the sequence (two copy of G-CSF with inteins and pelB) in pUC57 vector and then this sequence was cloned into the pET22b vector . The insertion of mentioned sequence into the pET22b vector was done by two 5' MscI and 3' XhoI restriction enzymes.
To start the fermentation, 50 mL baffled shake flasks containing 15 mL of the sterilized LB medium was inoculated with of E.coli BL21(DE3) and afterwards flasks were incubated in an incubator shaker at 37°C, agitated at 250 rpm. Cultures that supplemented with ampicillin for plasmid selection, were grown overnight at 37°C in liquid LB medium [14-16].
The growth of E.coli and production of G-CSF were tested by three media (4YT, LB and Optimized). 4YT consists of 32 g tryptone, 20 g yeast extract, 5g NaCl per liter and LB medium have glucose (0.4 g/L), tryptone (10.0 g/L), yeast extract (5.0 g/L) and NaCl (10.0 g/L). On the other hand, the composition of Optimized medium is 5.47 g/L glucose, 55.24 g/L yeast extract and 42.27 g/L peptone [17].
The recombinant cells were grown at 23°C , 30°C and 37°C in LB medium containing ampicillin at a concentration of 100 μg/ml. The isopropylthio-β-D-galactoside (IPTG) with amounts of 1mM was added when the cell concentration reached an optical density (600 nm) of about 0.5- 0.7. This IPTG was for inducing of the T7 promoter-operator. Then, the cells were harvested after 3h, 6h and 24 h incubation and the periplasmic fractions were prepared by two different osmotic shock procedures as described by Libby et al., [18] and Sockolosky [19].
The taking of cells and protein extraction were done at 2, 3 and 4 hours after induction time and afterwards proteins were visualized by SDS-PAGE. The dot blot method was used for ensuring of the expressed recombinant hG-CSF.
Periplasmic osmotic-shock fluid was obtained by Libby et al method (1987) and Sockolosky (2013). Briefly, the pellet was collected with centrifuging of 1.5 ml fermentation broth with OD600=1 unit at 15000×g for 5min. All the steps were carried out at 4°C. Pellets were re suspended in 15 μl of ice cold TES buffer (Tris-HCl 0.2 M, EDTA 0.5 M, Sucrose 0.5mM) pH 8.0, vigorously shaked every second minutes. 22.5 μl of ice cold double-distilled water was added and the incubation was continued for 30 min on ice. The cells were centrifuged at 16000 ×g for 20 min. Trichloroacetic acid was added to the supernatant up to 12% of the final volume. The mixture was centrifuged at 16000 ×g for 20 min [18].
According to another method, suspending of harvested cells was performed in a hypertonic solution of 30 Mm Tris, 20% w/v sucrose, 1 mM EDTA, pH 8 (25 mL) and incubation was carried out for 30 min at 4°C. Cells were centrifuged and the supernatant was collected. Re-suspending of cells was done in a hypotonic solution of 5 mM MgSO4 (25ml), incubation was continued for 30 min at 4°C and then followed by centrifugation. The supernatant from the hypertonic solution and the supernatant from the hypotonic solution were combined, centrifuged and cell debris was removed [19]). In sample buffer, pellet was dissolved and buffer was boiled for 5 min and saved for further analysis.
Periplasmic and cytoplasmic fractions Protein samples were separated by electrophoresis on a 10% (w/v) sodium dodecyl sulfate-polyacrylamide gel (SDS-PAGE) with Laemmli procedure [20]. and then gel was stained with Coomassie blue G-250. GCSF standard (PDgrastim from Pooyeshdarou pharmaceutical company) and protein samples were transferred into a nitrocellulose membrane and followed by stated for dot blotting as it was stated with anti-GCSF antibody [21] (Figure 4).
E.coli extracts and recombinant proteins were fractionated on a 12% SDS-PAGE and transferred to a nitrocellulose membrane. The blocking of membrane was carried out by 10 % (w/v) non-fat dried milk in phosphate buffered saline (0.05 % Tween 20 (PBS-T)) with shaking at room temperature, overnight. After three washes of 10 min with PBS-T, washing of membrane was done with shaking for 90 min with rabbit anti- GCSF (Santa cruz) in 5% non-fat dried milk-PBS-T (1:1,000), at room temperature. After three washes of 10 min with PBS-T, the incubation was continued for 60 min with goat anti-rabbit IgG HRP conjugate (Abcam) in 5 % non-fat dried milk- PBS-T at room temperature with shaking. After three more washes of 10 min with PBS-T, the developing of membrane was done by 4-Chloro-α-naphtol as western blotting detection reagent (Sigma) and then pink spots were appeared [2].
3mL LB medium that supplemented with ampicillin (100 µg/mL) was inoculated by a single colony and incubated at 37°C and then 0.5mL of this overnight culture was added into 50mL 4YT medium. At the final stage , this medium was severly shaken in 23°C for protein expression.
For cleaving of first G-CSF which its C-Terminal fused to intein2-CBD, DTT. was utilized and for cleaving of second GCSF that tagged to the CBD-intein1, pH and temperature shift was selected. Harvested cells after culture centrifugation, were suspended in buffer B1 (20mMTris, 500 mMNaCl, 1mM EDTA, pH 8.5) and were sonicated. At first, equilibration of chitin beads were done in buffer B1 (20mMTris, 500 mMNaCl, 1mM EDTA, pH 8.5). Then cells were lysed in buffer B1 by sonication and pellets were harvested , again pellets were dissolved in buffer B1 plus 2M urea. At the next stage, cell suspension is shaked slowly and after centrifuging in 14000 rpm , 2-8°C for 20 minutes , the clarified cell extract was applied for cleaving into the chitin column. After that, washing of the column was performed by buffer B1for removing of unbound proteins. After cleaving of first G-CSF, on-column cleavage of intein1 was induced by equilibrating of chitin beads in buffer B2 (20mMTris, 500 mMNaCl, 1mM EDTA, pH 6.5). Then, the cleavage reaction was continued overnight at room temperature. After 16 hours, column was washed by buffer B2 to remove any unbound protein species and hereafter washing was continued by buffer B1. At final stage, the cleavage of intein2 is induced by equilibrating of chitin beads in buffer B3 (20mMTris, 500 mMNaCl, 1mM EDTA ,40mM DTT, pH 8.5). and cleavage reaction was permitted to proceed overnight at room temperature. Later than 16 hours, column was washed by buffer B2 to remove any unbound protein species and hereafter washing was continued by buffer B1.
separation by electrophoresis on a 12% (w/v) SDSPAGE gel was done by Laemmli method for Protein samples, periplasmic and cytoplasmic fractions [20] and then gel was stained with Coomassie blue G-250 (Figure 6). Western blot: All of extracted proteins and GCSF standard were loaded on a 12 % SDS-PAGE and transferred to a PVDF membrane. The blocking of membrane was carried out by 10 % (w/v) non-fat dried milk in phosphate buffered saline (0.05 % Tween 20 (PBS-T)) with shaking at room temperature, overnight. After three washes of 10 min with PBS-T, washing of membrane was done with shaking for 90 min with rabbit anti- GCSF (Santa cruz) in 5 % non-fat dried milk-PBS-T (1:1,000), at room temperature. After three washes of 10 min with PBS-T, the incubation was continued for 60 min with goat anti-rabbit IgG HRP conjugate (Abcam) in 5 % non-fat dried milk-PBS-T at room temperature with shaking. After three more washes of 10 min with PBS-T, the developing of membrane was done by 4-Chloro-α-naphtol as western blotting detection reagent (Sigma) and then pink spots were appeared [2] ( Figure 7).
For studying of dimmers and monomers, SEC-HPLC was selected [16]. The HPLC system (Agilent , USA) was equipped with an SCL-10 Avp system controller, LC-10 ADvp pump, DGU-14A degasser,SIL-10ADvp auto-sampler, and an SPD-M10 Avp photodiodearray (PDA) detector. The detector was set at 214 nm and peak areas were integrated by computational analysis, using a Shimadzu Class VP® software program. Performing of experiments were done by column (300 × 7.8 mm I.D., with a particle size of 5 μmand pore size of 14.5 nm). The operation of HPLC system was at room temperature, using mobile phase of ammonium hydrogen carbonate and pHwas adjusted on 7 by the addition of phosphoric acid (10 N). The standard and sample were injected to the 20 μl, and all determinations were carried out in triplicate. Flow rate was 0.5 mL min-1. The Filgrastim standard was purchased from Pooyesh Darou pharmaceutical company.
For studying of biological activity, Male 7-8 week-old mice (19 -24g) were selected. Animals were kept under optimum conditions light, temperature and humidity, with food and water.
The animals were designated to sample, standard, and control groups randomly and identified by color code for assay (5 mice per group). Standard and test sample were prepared as the concentrations of 60μg mL-1, in phosphate buffered saline (PBS) with 0.1% bovine serum albumin (BSA).
Multiple injections of 0.5 mL rhG-CSF (standard or sample groups) were done to the animals in the treated groups, from day 1 to day 4. The administration in the same way was performed for control group with 0.1% BSA in PBS. The peripheral blood was collected six hours after the last injection. Smears were prepared on glass slides and staining was done by the May- Grunewald-Giemsa method, and the white cells and total number of neutrophils were counted. The results (% of neutrophils) are presented as the mean ± S.D. of 6 animals. Statistical comparison of data was performed by one-way analysis of variance (ANOVA) followed by Bonferroni's post-test, by means of GraphPad Prism Program (Version 4.0). P-values less than 0.05 (P < 0.05 or less) were considered significant.
For determining the possible differences in the biological activity between the standard and the sample groups, the Test described by Finney [22] was adopted, through the parallel line method (3 × 3), using a PLA 1.2 program (Stegmann Systemberatung, Rodgau, Germany). Analysis of variance was performed for each assay, and the assumption of linearity and parallelism of the log dose log response lines was tested (P < 0.05). Statistical weights were computed as the reciprocal of the variance of the log potency. Estimates of log potency were examined for heterogeneity using a Χ² test (P = 0.05) and were combined as weight geometric means of homogeneous estimates (P > 0.05).
The effect of three different types of basal medium (4YT, LB and Optimized) on growth of E.coli and production of G-CSF was shown that the highest growth of E.coli and production of G-CSF was belonged to 4YT and Optimized media. The slightly inhibition in growth and great reduction in the production of G-CSF was observed in LB medium. Also, Sockolosky method was the best method of osmotic shock for periplasmic protein isolation (2013).
Observation of protein bands of approximately 43.7kDa and 53.7kDa corresponding to the molecular weights of GCSF-intein2 and intein1-GCSF was done, after induction (Figure 1, shown by arrows). We had best expression, whenever the IPTG concentration was received to 1mM and 23°C was induction temperature.
Therefore, induction with 1mM IPTG for 24 h at 23°C was used for further experiments as optimized conditions for expression of GCSF-intein2 and intein1- GCSF.
Dot blot analysis showed that production of human GCSF in 4YT medium is good and protein was detectable in both cell lysate and medium [Figure 2]. Afterwards, producing of human GCSF in E.coli cultured in 4YT medium was shown by western blot and proteins was detectable in both cell lysate and medium [Figure 3].
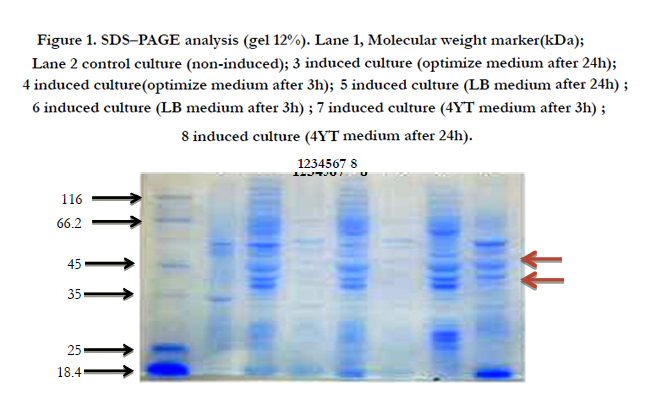
Figure 1. SDS–PAGE analysis (gel 12%). Lane 1, Molecular weight marker(kDa); Lane 2 control culture (non-induced); 3 induced culture (optimize medium after 24h); 4 induced culture(optimize medium after 3h); 5 induced culture (LB medium after 24h) ; 6 induced culture (LB medium after 3h) ; 7 induced culture (4YT medium after 3h) ; 8 induced culture (4YT medium after 24h).


Figure 2. Dot blot analysis of E. coli expressed rhGCSF in 4YT medium against of standard 1,2: Standard dilutions (200 μg/ml , 20 μg/ml , 2 μg/ml , 0.2 μg/ml), 3,4,5: 4YT medium dilutions (200 μg/ml , 20 μg/ml , 2 μg/ml , 0.2 μg/ml) and 6: non-induced culture.
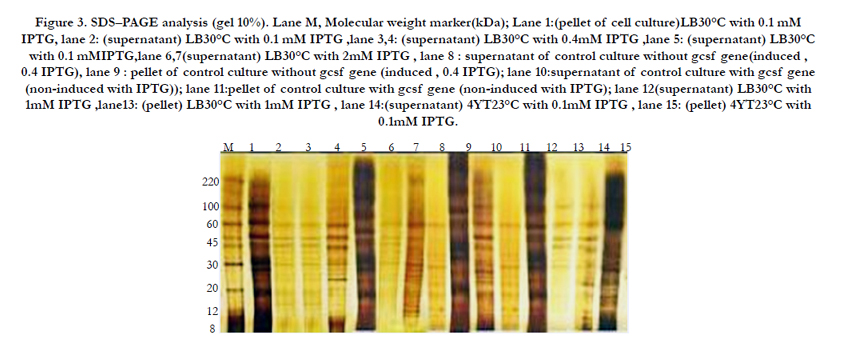

Figure 3. SDS–PAGE analysis (gel 10%). Lane M, Molecular weight marker(kDa); Lane 1:(pellet of cell culture)LB30°C with 0.1 mM IPTG, lane 2: (supernatant) LB30°C with 0.1 mM IPTG ,lane 3,4: (supernatant) LB30°C with 0.4mM IPTG ,lane 5: (supernatant) LB30°C with 0.1 mMIPTG,lane 6,7(supernatant) LB30°C with 2mM IPTG , lane 8 : supernatant of control culture without gcsf gene(induced , 0.4 IPTG), lane 9 : pellet of control culture without gcsf gene (induced , 0.4 IPTG); lane 10:supernatant of control culture with gcsf gene (non-induced with IPTG)); lane 11:pellet of control culture with gcsf gene (non-induced with IPTG); lane 12(supernatant) LB30°C with 1mM IPTG ,lane13: (pellet) LB30°C with 1mM IPTG , lane 14:(supernatant) 4YT23°C with 0.1mM IPTG , lane 15: (pellet) 4YT23°C with 0.1mM IPTG.
For determination of rhG-CSF biological potency, we used an in vivo model of healthy mice. The percentage of neutrophils in serum was estimated 6h after the last rhG-CSF injection. The figure 8 demonstrate that treatment with rh-GCSF in the sample group caused an important increase of neutrophil numbers in compared to the control group. The neutrophil numbers in the reference group were remarkably different from that observed in the sample group compared with the same doses of rhGCSF. Statistical analysis by the Finney [22] exhibited a biological activity of 109.4% for the rhG-CSF in the sample group. It appears that the assay of Filgrastim (rhG-CSF) itself is not described in any Pharmacopoeia, although values ranging from 90 to 110% have been suggested for biological drugs, according to the European Pharmacopoeia [24]. The data obviously show that the mixture of experimental procedures made the rhG-CSF that are comparable with the standard rhG-CSF. It is tempting to infer that on a large scale up production with these methods can generate rhG-CSF with significant amount and high biological activity. At first, the human G-CSF formally was used for leukopenia treatment in US for 1991. At present, the hG-CSF is widely utilized and effectively is useful as haematopoietic growth factor. However, protein secretion is a complex and incompletely understood process, therefore. Despite of using of signal peptides and unsuccessful secretion of proteins in periplasmic space, the secretion of a variety of heterologous proteins has been done by pelB signal peptide. Our prokaryotic expression vector pET22b that contained Intein1- GCSF - Spacer - GCSF - intein2 sequence, and achieved good expression of the hG-CSF in E.coli, and this expression with SDS-PAGE, dot blot and western blot was shown (Figure 5).
The results showed that in 4YT medium after 24 hours induction with 1mM IPTG, the highest periplasmic expression level was occured, that is with agreement with previous experiments, outlined in Figure 3. It seems that if culture medium, temperature and the other important factors in expression level is optimized, hG-CSF into periplasmic space has effective production. It is also indicated that pelB signal peptide may be useful for periplasmic production of other recombinant proteins in E.coli. There have also been some studies showing that the use of signal sequences such as pelB do not result in the complete secretion of mature proteins in E.coli [18, 19]. In this report, G-CSF protein was expressed in the form of inclusion bodies in E.coli and was successfully in vitro refolded through urea denaturation. The refolding recovery of the fusion product reaches about 69%. About 60 μg/ml protein was achieved by single-step purification on chitin beads column,without any chemicals or protease treatment.In conclusion, production of proteins in periplasm help the pharmaceutical companies to produce bioproducts as correct and soluble form and likewise, intein-mediated purification scheme can help to these companies for economic purification of recombinant proteins.

Figure 4. SDS–PAGE analysis (gel 10%). Lane M, Molecular weight marker(kDa); Lane 1:(pellet) 4YT23°C with 1mM IPTG, lane 2: (supernatant) 4YT23°C with 1mM IPTG,lane 3: (pellet) 4YT23°C with 0.4mM IPTG, lane 4:(supernatant) 4YT23°C with 0.4mM IPTG , lane 5: (pellet) 4YT23°C with 2 mM IPTG , lane 6: (supernatant) 4YT23°C with 2 mM IPTG , lane 7: (pellet) LB30°C with 2 mM IPTG , lane 8: (pellet) Optimized medium 37°C with 2 mM IPTG , lane 9: (supernatant) Optimized medium37°C with 2 mM IPTG , lane 10: (pellet) Optimized medium37°C with 1mM IPTG , lane 11: pellet of control culture with gcsf gene (non-induced with IPTG , 23°C) , lane 12: supernatant of control culture with gcsf gene (non-induced with IPTG , 23°C) , lane 13: pellet of control culture without gcsf gene (induced , 0.4 IPTG , 23°C) , lane 14: supernatant of control culture without gcsf gene (induced , 0.4 IPTG , 23°C)
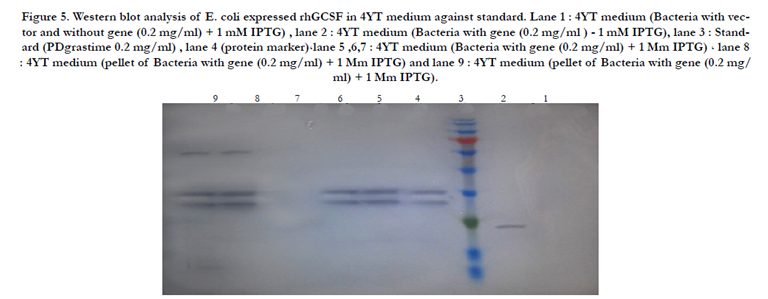
Figure 5. Western blot analysis of E. coli expressed rhGCSF in 4YT medium against standard. Lane 1 : 4YT medium (Bacteria with vector and without gene (0.2 mg/ml) + 1 mM IPTG) , lane 2 : 4YT medium (Bacteria with gene (0.2 mg/ml ) - 1 mM IPTG), lane 3 : Standard (PDgrastime 0.2 mg/ml) , lane 4 (protein marker)،lane 5 ,6,7 : 4YT medium (Bacteria with gene (0.2 mg/ml) + 1 Mm IPTG) ، lane 8 : 4YT medium (pellet of Bacteria with gene (0.2 mg/ml) + 1 Mm IPTG) and lane 9 : 4YT medium (pellet of Bacteria with gene (0.2 mg/ ml) + 1 Mm IPTG).

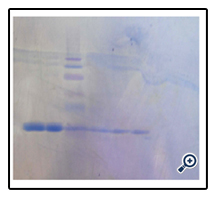
Figure 6. SDS–PAGE analysis (gel 12%). Lane 1,2 :Standard GCSF (PDgrastim), Lane 3 :Molecular weight marker (kDa); Lane 4, 5,6 : purified GCSF.
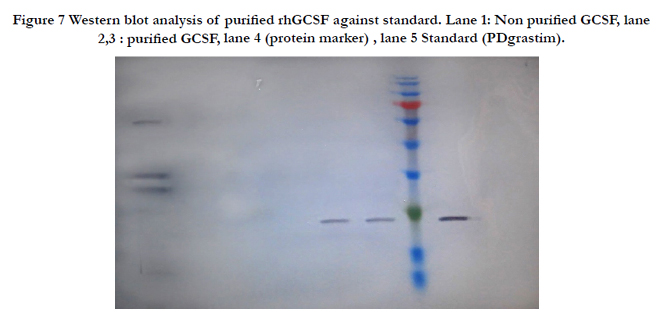
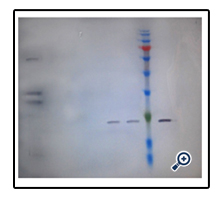
Figure 7 Western blot analysis of purified rhGCSF against standard. Lane 1: Non purified GCSF, lane 2,3 : purified GCSF, lane 4 (protein marker) , lane 5 Standard (PDgrastim).
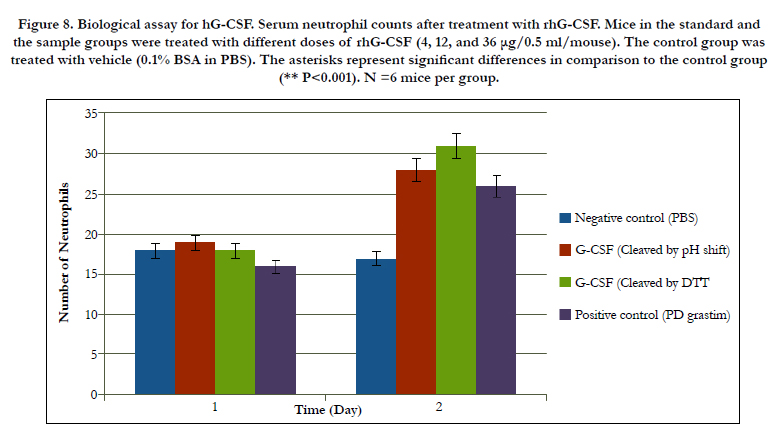
Figure 8. Biological assay for hG-CSF. Serum neutrophil counts after treatment with rhG-CSF. Mice in the standard and the sample groups were treated with different doses of rhG-CSF (4, 12, and 36 μg/0.5 ml/mouse). The control group was treated with vehicle (0.1% BSA in PBS). The asterisks represent significant differences in comparison to the control group (** P<0.001). N =6 mice per group.
Acknowledgments & Declarations
This study was supported by University of Isfahan and Exir pharmaceutical company.
References
- Marco A D (2009) Strategies for successful recombinant expression of disulfide bond-dependent proteins in Escherichia coli. Microbial Cell Factories.8 (26): 1-18.
- Fallah M J, Akbari B, Saeedinia A R, Karimi, Vaez M, et al. (2015) Overexpression of recombinant human granulocyte colony-stimulating factor in E. coli. Ir J Med Sci. 28(3): 131-134.
- Shahali, M, Yakhchali, B, Zomorodipour, A, & Seyedena, S. Y. (2005) Expression and Secretion of Human Granulocyte Macrophage-Colony Stimulating Factor Using Escherichia coli Enterotoxin I Signal Sequence. J Sci. 16(4): 327-332.
- Jeong KJ, Lee, S Y (2001) Secretory production of human granulocyte colony-stimulating factor in Escherichia coli. Pro Exp Pur. 23 (2): 311–318.
- Jin H, Cantin G T, Maki S, Chew L C, Resnick S M, et al. (2011) Soluble periplasmic production of human granulocyte colony-stimulating factor(GCSF) in Pseudomonas fluorescens. ProExpPur. 78 (1): 69-77.
- Choi J H, Jeong K J, Kim S C, Lee S Y (2000) Efficient secretory production of alkaline phosphatase by high cell density culture of recombinant Escherichia coli using the Bacillus sp. endoxylanase signal sequence. Appl Microbiol Biotechnol. 53 (6): 640-645.
- Thomas J, Liu F, Link DC (2002) Mechanisms of mobilization of hematopoietic progenitors with granulocyte colony-stimulating factor. CurrOpHematol. 9 (3): 183–9.
- Pessach I, Shimoni A, Nagler A (2013) Granulocyte-colony stimulating factor for hematopoietic stem cell donation from healthy female donors during pregnancy and lactation: what do we know?. Human Reproduction Update 19 (3): 259–67.
- NishiiK , Xing X-H, Shiragami N, Unno H (1991) Production of r G-CSF by CHO cell in aggregate microbeads culture. Cytotechnology. (7): 25-32.
- Bahrami A, Shoja Alsadati S A, Khalilzadeh Rasoul, Saeidi Nia A R, Vasheghani Faraahani E, et al. (2005) Production of recombinant human granulocyte-colony stimulating factor by Pichia pastoris. IrJBiotechnol. 5(3): 162-169.
- Chien S F (2010) Cloning and expression of bioactive human granulocyte colony stimulating factor in Pichia pastoris. J Chin Chem Soc. 57 (4B):850-856.
- Guo CH, Lia Z, Shia Y, Xub M, Wise JG, et al. (2004) Intein-mediated fusion expression, high efficient refolding,and one-step purification of gelonintoxin. ProExpPur. 37 (2): 361–367.
- Babu K S, Muthukumaran T, Antony A, Samuel S D P S, Balamurali M, et al. (2009) Single step intein-mediated purification of hGMCSF expressed in salt-inducible E. coli. BiotechnolLett . 31 (5): 659–664.
- Do B H, Ryu H B , Hoang PH, Koo B K, Choe H (2014) Soluble prokaryotic over expression and purification of bioactive human granulocyte colonystimulating factor by maltose binding protein and protein disulfide isomerase.PLOS ONE. 9 (3): e89906.
- Nagata S, Tsuchiya M, Asano S, Kaziro Y, Yamazaki T, et al. (1986) Molecular cloning and expression of cDNA for human granulocyte colony-stimulating factor". Nature 319 (6052): 415–8.
- Vanz A LS , Renard G , Palma MS, Chies J M, Dalmora S L, et al. (2008) Human granulocyte colony stimulating factor (hG-CSF): cloning, over expression,purification and characterization. Microbial Cell Factories. 7 (13):1-12.
- Tan J S, Ramanan R N, Azaman S N A, Ling T C, Shuhaimi M et al. (2009) Enhanced Interferon-α2b Production in Periplasmic Space of Escherichia coli through Medium Optimization using Response Surface Method.Biotechnol J. (3): 117-124.
- Libby R, Braedt G, Kronheyim S, March CJ, Urdal DL, et al. (1987) Expression and purification of native human granulocyte-macrophage colonystimulating factor from an Escherichia coli secretion vector. DNA. 6 (3):221-229.
- Sockolosky JT, Szoka FC (2013) Periplasmic production via the pET expression system of soluble, bioactive human growth hormone. ProExpPur.87 (2): 129–135.
- Lammli UK (1970) Cleavage of structural protein during the assembly of head of bacteriophage T4. Nature. 15 (227): 680-685.
- Rezaei M and Zarkesh-Esfahani S H (2012) Optimization of production of recombinant human growth hormone in Escherichia coli. J Res Med Sci. 17(7): 681–685.
- Finney JD (1978) Statistical methods in biological assay. Charles Griffin London; 1978. 21 (7): 689–690.
- Dalmora SL, Massiero S, Oliveira P, Sangoi M, Brum L (2006) Validation of an RP-HPLC method and assessment of rhG-CSF in pharmaceutical formulation by liquid chromatography and biological assay. Journal of liquid chromatography & related technologies, 29 (12): 1753-1767.
- Gaines D, Tydeman M (1980) Iterative weighted regression analysis of logit responses: a computer program for the analysis of bioassay and immunoassays.Comput Programs Biomed,15 (1): 13-21.





