Cloning and functional expression of zeta-carotene desaturase, a novel carotenoid biosynthesis gene from Ficus carica.
J.M. Araya-Garay1*, L. Feijoo-Siota2, P. Veiga-Crespo2,3, A. Sanchez-Perez4, T. Gonzalez Villa2,3
1 Interdisciplinary Center for Aquaculture Research (INCAR), University of Concepcion, Concepcion. Chile.
2 Department of Microbiology, University of Santiago de Compostela. Spain.
3 School of Biotechnology. University of Santiago de Compostela. 15782 Santiago de Compostela, Spain.
4 Discipline of Physiology and Bosch Institute, University of Sydney, NSW 2006, Australia.
*Corresponding Author
J.M. Araya-Garay,
Laboratory of Biotechnology and Aquatic Genomics Interdisciplinary Center for Aquaculture Research (INCAR).
University of Concepcion,
Concepción, Chile.
Tel: +56 988450331
E-mail: josemiguel.araya@gmail.com
Article Type: Research Article
Received: December 15, 2013; Accepted: January 25, 2014; Published: January 27, 2014
Citation: J.M. Araya-Garay, et al (2014) Cloning and functional expression of zeta-carotene desaturase, a novel carotenoid biosynthesis gene from Ficus carica. Int J Microbiol Adv Immunol. 2(1), 32-40. doi: dx.doi.org/10.19070/2329-9967-140006
Copyright: J.M. Araya-Garay © 2014. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
Carotene desaturation, an essential step in the carotenoid biosynthesis pathway, is catalyzed by two enzymes, phytoene desaturase (PDS) and ζ-carotene desaturase (zeta carotene desaturase, ZDS). Here we describe cloning and E. Coli expression of zds-Fc, a novel Ficus carica ζ-carotene desaturase catalyzing dehydrogenation of ζ-carotene into neurosporene and finally lycopene. The ζ-carotene desaturase (ZDS) gene was amplified from the fig tree by rapid amplification of cDNA ends (RACE) and spanned a 1746 bp open reading frame (ORF), encoding a protein of 582 amino acid residues with a predicted molecular weight of 64kD. The N-terminal region of this polypeptide contained a putative transit sequence for plastid targeting. By phylogenetic and sequence analyses, zds-Fc showed high homology with previously described ζ-carotene desaturases from higher plant species (Al-Babili et al. 1998; Cong et al. 2009; Matthews et al. 2003; Yan et al. 2011). Additionally, sequence analysis revealed a high degree of conservation among plant ZDSs. The deduced ZDS protein, designated FcZDS, also contains an N-terminus dinucleotide-binding, followed by a conserved region identified in other carotene desaturase sequences. These data, taken together, confirm our cloned zds-Fc as an integral part of the ZDS family of proteins.
2.Introduction
3.Materials and Methods
3.1.Materials
3.2.Cloning of Ficus carica ζ-carotene desaturase cDNA (zds-Fc)
3.3.RACE-PCR
3.4.Bacterial transformation and growth
3.5.Protein electrophoresis (SDS-PAGE)
3.6.Bioinformatic analyses
3.7.HPLC analysis
3.8.Application of sELISA
3.9.Cutoff value estimation
3.10.Evaluation of sELISA
3.11.Characterization of H. pylori sonicate whole cell protein by SDS-PAGE
3.12.Ethical clearance
4.Results
4.1.Requirement for acceptance of ELISA plates
4.2.Rabbit anti- H. pylori serum valuation
4.3.Phylogenetic analysis of plant ZDS proteins
4.4.Carotenoid production and ZDS-Fc enzymatic activity determination
5.Discussion
6.Acknowledgements
7.References
Keywords
Ficus Carica; Zeta-Carotene Desaturase; Cdna; Zeta-Carotene; Neurosporene; Lycopene.
Introduction
Carotenoids are pigments synthetized by plants, fungi, bacteria, and algae with the main function of protecting them from the action of singlet oxygen and other radicals (Krinsky 1989). In plants, carotenoids are either primary or secondary metabolites. As primary metabolites, carotenoids can function as regulators of plant growth and development, as accessory pigments in photosynthesis, as photoprotectors preventing photo- oxidative damage, or as precursors to the hormone abscisic acid (ABA) (Hirschberg 2001). They are also responsible for the color of fruits and flowers, generating distinct yellow, orange, and red colors, thus substantially contributing to plant-animal communication (Hirschberg 2001; DellaPenna and Pogson 2006). In addition, the colors of many carotenoid-accumulating fruits and flowers also increase their appeal and hence their economic value (Fraser and Bramley 2004; Rodríguez Concepcion 2006). These pigments also play an important protective role, in human and animal diets, as antioxidants.
The main carotenoid metabolic pathway is well known and may be shared by most of the carotenogenic species (Sandmann 2002). The first step in this pathway is the condensation of two molecules of geranyl geranyl pyrophosphate (GGPP) to originate the first true carotenoid C40 molecule, phyotene (Park et al. 2002). This two-step reaction is catalyzed by a single soluble enzyme: phytoene synthase (PSY) (Fig. 1). Two sequential desaturations of phytoene result in the formation of the first phytofluene, followed by ζ-carotene. Both of these reactions are catalyzed by phytoene desaturase (PDS). Two additional desaturations, catalyzed by ζ-carotene desaturase (ZDS), give rise to first neurosporene and finally lycopene, the symmetrical red carotenoid pigment (Sandmann 1994) (Fig. 1).

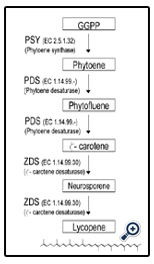
Figure 1: Schematic diagram of the early steps in the carotenoid biosynthetic pathway in plants, from geranyl geranyl pyrophosphate (GGPP) to lycopene, and the enzymes catalyzing those reactions.
Since 1990, when Misawa (Misawa et al. 1990) cloned the carotenoid biosynthesis gene cluster from Erwinia uredovora, many carotenoid biosynthetic genes have been identified in plants and other organisms (Cong el al. 2009). We have previously reported (Araya-Garay et al. 2011) the production of β-carotene by expression of recombinant Ficus carica lycopene beta-cyclase in E. Coli. Here, we describe cloning and Escherichia coli expression of zds-Fc, a novel Ficus carica ζ-carotene desaturase catalyzing dehydrogenation of ζ-carotene into neurosporene and finally lycopene. The E. Coli culture containing zds-Fc was able to convert ζ-carotene and neurosporene, two substrates of ζ-carotene desaturase, into neurosporene and lycopene, or lycopene, respectively; thus confirming, in vivo, the enzymatic activity of the recombinant ZDS we produced. The F. carica enzyme also shared the characteristic properties of other plant ZDSs, and this allowed us to construct a phylogenetic tree illustrating the evolutionary relationship between ZDS and other published carotenoid zeta carotene desaturases (ZDSs) from higher plants.
Materials and Methods
Plasmids pACCRT-EBI (Misawa and Shimada 1998) and pACCRT-EBR (Takaichi et al. 1996), encoding Erwinia uredovora crtE (GGPP synthase), crtB (phytoene synthase) and crtI (phytoene desaturase), and encoding Erwinia uredovora crtE, crtB plus Rodococcus crtR, respectively, were a present from Prof. Misawa (Research Institute for Bioresources and Biotechnology, Ishikawa Prefectural University, Japan). Whereas plasmid pACCRT-EBP (Linden et al. 1993), encoding Erwinia uredovora crtE, crtB, plus Synechococcus crtP was kindly donated by Prof. Gerhard Sandmann (J. W. Goethe Universität, Frankfurt, Germany). Plasmids pCR Blunt II Topo and pUC19, as well as E. Coli strains TOP10 and BL21 (DE3) and Zero Blunt TOPO PCR Cloning Kit, were from Invitrogen (Carlsbad, CA, USA). The expression plasmid pET21a was purchased from Novagen, (Cambridge, UK). RNeasy Plant Mini Kit was from Qiagen (Valencia, CA, USA). The RT-PCR AMV kit was obtained from Roche Applied Science (Indianapolis, IN, USA). Accuzyme DNA polymerase was from Bioline (Taunton, MA, USA).
Total RNA was isolated, using the RNeasy Plant Mini Kit, from 50 mg of fresh Ficus carica leaf, following the instructions recommended by the manufacturer. RTPCR was carried out using the RT-PCR AMV kit and degenerate primers ZDS DegF and ZDS DegR (Table 1). These oligonucleotide primers were designed using Primer Premier 5.0 (Biosoft International), according to the conserved motifs found on other published zeta-carotene desaturase sequences. Amplification of zds-Fc conserved internal DNA fragment was achieved by PCR, using the above-described oligonucleotide primers and 2 μL of first-strand cDNA. The reaction mixture contained polymerase buffer, 0.2 mM of each primer, 1.5 mM MgCl2, 0.2 mM of each deoxynucleotide, and 1 unit of Accuzyme DNA polymerase, following program: 94°C for 2 min, 35 cycles of 94°C for 45 s; 49°C for 1 min and 72°C for 2 min; and a final extension at 72°C for 2 min. The PCR product obtained was subcloned into pCR Blunt II TOPO, using the Zero Blunt TOPO PCR Cloning Kit. All DNA constructs were checked by sequence analysis.


*Note: F, forward; R, reverse; ORF, open reading frame.
Table 1: PCR primers used in this study, their oligonucleotide sequence is shown 5’ to 3’, and the purpose the primers were generated for.
The 5´and 3´ ends of zds-Fc were obtained by RACE (rapid amplification of cDNA ends), using the 5´/3´ RACE kit, 2nd Generation (Roche Applied Science), primers ZDS-Fc1, ZDS-Fc2, ZDS-Fc3, ZDS-Fc4 and ZDS-Fc5 (Table 1), and 1 unit of Accuzyme DNA Polymerase. ZDS-Fc1 was the primer used for first strand cDNA synthesis, whereas the missing 5´ region of the gene was amplified with ZDS-Fc2 and Oligo dT-Anchor Primer, and incubated as follows: 1 cycle at 94°C for 2 min; 10 cycles of 94°C for 15 s, 55°C for 30 s and 72°C for 40 s; 25 cycles of 94°C for 15 s, 55°C for 30 s and 72°C for 40 s, increasing the elongation time by 20 s for each successive cycle (i.e. elongation time of cycle no.11 is 40 s, of cycle no. 12 is 60 s, of cycle no. 13 is 80 s etc.), and a final extension at 72°C for 7 min.
The PCR-amplified DNA product was then used as a template (dilution 1:20) for a subsequent PCR, with ZDS-Fc3 and PCR Anchor Primer as primers. This second PCR was incubated as described above, with the exception of the primer annealing step, carried out at 60°C.
The 3´end of zds-Fc gene was amplified in two sequential PCR reactions, using ZDS-Fc4 and ZDS-Fc5, respectively, and the PCR Anchor as primers. The DNA sequence of the PCR-amplified fragments was confirmed by sequence analysis.
PCR primers ZDS-EcoRI F and ZDS-XhoI R were designed to amplify the complete zds-Fc coding region (Table 1), using the above described zds-Fc DNA fragments as template. These primers contained EcoRI and XhoI restriction sites at their 5’ and 3´ ends, respectively (in bold in Table 1). The PCR conditions were: 1 cycle at 95°C for 3 min; 35 cycles of 95°C for 10 s, 57°C for 10 s and 72°C for 2 min ; and a final extension at 72°C for 10 min. The resulting PCR product was cloned into a pET21a vector, using the restriction sites present in the primers, thus producing the expression plasmid pET21-zdsFc (Table 2).
Transformation of plasmid pACCRT-EBI into E.Coli BL21 (DE3) generated E.Coli BL-pEBI (Table 2), a bacterial strain expressing the enzymes GGPP synthase, phytoene synthase and phytoene desaturase, from Erwinia uredovora, and hence capable of producing lycopene. This recombinant E.Coli was used as positive control in our zeta-carotene desaturase enzymatic assay (see below). Transformation of plasmid pACCRT-EBP into E.Coli BL21 (DE3) resulted in E.Coli BL-EBP (Table 2), a bacterial strain expressing Eu-crtE, Eu-crtB and Sc-crtP. E.Coli BLpEBR (Table 2) was generated by transforming E.Coli BL21 (DE3) with plasmid pACCRT-EBR, encoding Eu-crtE, Eu-crtB and Rc-crtR, and thus capable of producing neurosporene. These recombinant E.Coli strains were used for heterologous complementation assays (see below). All recombinant E.Coli strains were grown in LB medium (1 % tryptone, 0.5 % yeast extract, 1 % NaCl) for 48 h at 28°C, with agitation at 180 rpm, in the dark to maximize carotenoid production. Chloramphenicol (34 μg/ml) was used for selection of E.Coli BL-EBI, BL-EBP and BL-EBR transformants. The culture broth was supplemented with 2 % agarose for growth on solid medium. Plasmid pET21a, containing the complete coding area of the zds-Fc gene (pET21-zdsFc), was transformed into E.Coli BL21 (DE3) to produce E.Coli BL-zdsFc (Table 2). E.Coli BL-zdsFc was grown in LB medium for 48 h at 28°C, in the presence of ampicillin (50 μg/ml), 1 mM IPTG (Isopropyl-β-D thiogalactopyranoside) was added at the end of the logarithmic growth phase to induce ZDS protein expression.
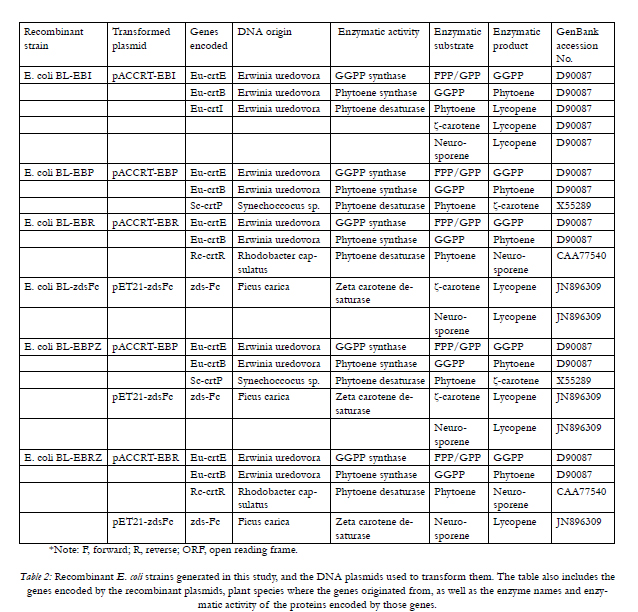
*Note: F, forward; R, reverse; ORF, open reading frame.
Table 2: Recombinant E. coli strains generated in this study, and the DNA plasmids used to transform them. The table also includes the genes encoded by the recombinant plasmids, plant species where the genes originated from, as well as the enzyme names and enzymatic activity of the proteins encoded by those genes
For protein analysis, recombinant E coli BL21 (DE3) cells, containing either pET21a or pET21-zdsFc, were grown at 37°C, with orbital shaking (180 rpm), until they reached the exponential growth phase. The cultures were then induced for recombinant protein expression, by addition of 1mM IPTG, and incubated for a further four hours before sample collection and centrifugation, as recommended by the pET21a system (Novagen). Prior to polyacrylamide gel electrophoresis, the bacterial cells were solubilized, at 85°C for 3 min, in a buffer containing 2.5% (w/v) SDS, 125 mM dithiothreitol, 25% (v/v) glycerol, and 112.5 mM Tris-HCl, pH 6.8. The protein samples were then loaded onto a 12% polyacrylamide gel, on a Mini Protean II cell (Bio-Rad, Hercules, CA), as described previously (Laemmli 1970). The gel was run, at 120V for 1 h, using Tris-Glycine buffer (25 mM Tris, 192 mM Glycine, 0.1% SDS, pH: 8.3). The molecular weight of the proteins was estimated by comparing their gel migration patterns to those of Precision Plus Protein Standard (BioRad), using the Quantity One software (Bio-Rad).
To construct the phylogenetic tree (Figure 2), the inferred fig tree (Ficus carica) ZDS amino acid sequence was compared to 22 homologous amino acid sequences from three citrus trees (Citrus sinensis, C. unshiu and C. maxima), physic nut tree (Jatropha curcas), pawpaw (Carica papaya), strawberry (Fragaria x ananassa), two apple trees (Malus x domestica), peppers (Capsicum annuum), tomato (Solanum lycopersicum), sweet potato (Ipomea sp.), gentian (Gentiana lutea), chrysanthemum (Chrysanthemum x morifolium), marigold (Tagetes erecta), two wild carrots (Daucus carota), daffodil (Narcissus tazetta var. Chinensis), orchid (Oncidium Gower Ramsey), cress (Arabidopsis thaliana), turnip (Brassica rapa), sorghum (Sorghum bicolour), wheat (Triticum aestivum), and green algae (Dunaliella salina).
Vector NTI Advance 11 software (Invitrogen) and BioEdit Sequence Alignment Editor version 7.0.5.3, were used to analyze the nucleotide and deduced amino acid sequences, and for sequence alignment, respectively. The NCBI database was searched for plant ZDS sequences using the BLAST software (Altschul et al. 1990). The ChloroP 1.1 Prediction Server program (Emanuelsson et al. 1999) and TargetP 1.1 Server (Emanuelsson et al. 2000; Nielsen et al. 1997) were used to identify the ZDS signal/sorting peptide and for predicting its cleavage site. Phylogenetic and molecular evolutionary analyses were conducted using MEGA, version 5.0, package program (Tamura et al 2011). Data were analyzed by the neighbor-joining method (Saitou and Nei 1987). The reliability of the neighbor-joining tree was estimated by calculating bootstrap confidence limits (BCL) (Saitou and Nei 1987), based on 1000 replicates. The evolutionary distances were computed using the Jones-Taylor-Thornton method (Nei and Kumar 2000) and are in the units of the number of base substitutions per site. The rate variation among sites was modeled with a Gamma distribution (shape parameter = 1). The Gen Bank accession numbers of the amino acid sequences used in the analysis are shown in Figure 2. Protein sorting was predicted by PSORT, a web server (Nakai and Horton 1999) for analyzing and predicting protein-sorting signals, from the Institute for Molecular and Cellular Biology (Osaka, Japan). Finally, PSIpred v3.0 (Jones 1999) was used for hydrophobicity and protein secondary structure predictions.
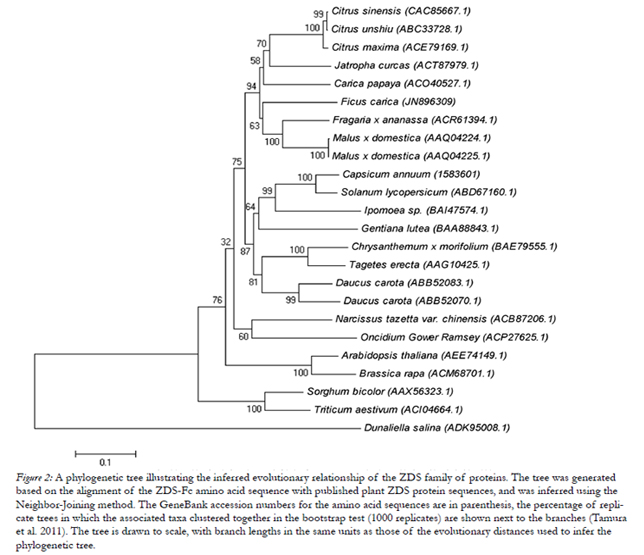
Figure 2: A phylogenetic tree illustrating the inferred evolutionary relationship of the ZDS family of proteins. The tree was generated based on the alignment of the ZDS-Fc amino acid sequence with published plant ZDS protein sequences, and was inferred using the Neighbor-Joining method. The GeneBank accession numbers for the amino acid sequences are in parenthesis, the percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1000 replicates) are shown next to the branches (Tamura et al. 2011). The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree.
For HPLC analysis, E. Coli BL-EBPZ and BL-EBRZ (Table 2) were grown overnight in LB broth, supplemented with chloramphenicol (34μg/ml) and ampicillin (50μg/ml), at 37°C with agitation (200 rpm). Recombinant protein expression was then induced, by IPTG addition (1mM), and the cultures incubated, in the dark, at 28°C for 48h with orbital shaking (200 rpm) After incubation, the cells were centrifuged (4000xg for 5 min), and the bacterial pellet washed twice with deionized water, re-suspended in acetone (Sigma) and homogenized by vortexing (10 min at 4°C). After a new centrifugation (13000xg, 2 min, 4°C), the cell supernatant was collected, dried down under N2 flow, and stored at -80°C for high performance liquid chromatography (HPLC) analysis. All sample manipulations were carried out on ice, under dim light, to prevent photodegradation, isomerization, or other structural changes in the recombinant terpenoids. For HPLC analysis, the dry residues were suspended in 2 ml of chlorophorm:metanol:acetone (3:2:1, v/v) and filtered through a 0.22 μm polycarbonate filter. The HPLC device was equipped with a photodiode array detector,set to scan from 250 to 540 nm throughout the elution procedure, and controlled by the Empower2 software program. Twenty μl of the terpenoid samples were loaded onto a C30 carotenoid column (250mm x 4,6mm, 5μm; YMC Europa) and the flow rate was maintained at 1ml/min. The mobile phase was: A, methyl tert-butil ether; B, water; and C, methanol. The linear ternary gradient elution program was performed as follows: Initially A-B-C (5:5:90); followed by 0-12 min, A-B-C (5:0:95); 12-20 min, A-B-C (14:0:86); 20- 30 min, A-B-C (25:0:75); 30-50 min, A-B-C (50:0:50); 50-70 min, A-B-C (75:0:25); and finally back to A-B-C (5:5:90) for column re-equilibration at 23°C. A Maxplot chromatogram, which plots each carotenoid peak and its corresponding maximum absorbance wavelength, was obtained for each HPLC sample; and the recombinant proteins were identified by comparing their HPLC retention profiles to those of standards run in the same HPLC conditions, or to published data. The lycopene standard was obtained from Sigma- Aldrich (Madrid, Spain), and used as previously described (Melendez Martinez et al. 2003; Rodrigo et al. 2004), whereas ζ-carotene and neurosporene were produced by the recombinant E. Coli cultures BL-EBP and BL-EBR, respectively. Quantitative analyses were carried out by comparing the Maxplot chromatogram peak areas of the carotenoid samples to a calibration curve constructed with the lycopene standard.
Zeta-carotene desaturase enzymatic activity was determined by functional complementation assays on E. Coli strains BL-pEBP and BL-pEBR. These strains showed a light yellow and dark yellow coloration, due to ζ-carotene and neurosporene accumulation, respectively. Both E. Coli BL-pEBP and BL-pEBR were -transformed with the plasmid pET21-zdsFc, to generate E. Coli BL-EBPZ and E. Coli BL-EBRZ, respectively. Transformed cells were plated onto LB agar plates containing chloramphenicol (34μg/ml) and ampicillin (50μg/ml). These plates also contained 10μl of a 100 mM aqueous solution of IPTG, added one hour before the cells were plated. The cells were incubated at 28ºC, for 48 h in the darkness, to maximize terpenoid production. After incubation, the desaturase enzymatic activity on the zeta-carotene and neurosporene substrates produced by the E. Coli strains, was identified by the pigments present on the recombinant colonies, and confirmed by HPLC analysis. The negative control for our assay was an E. Coli BL21 (DE3) containing the plasmid pET21a alone, whereas recombinant E. Coli BL-pEBI was used as the positive control.
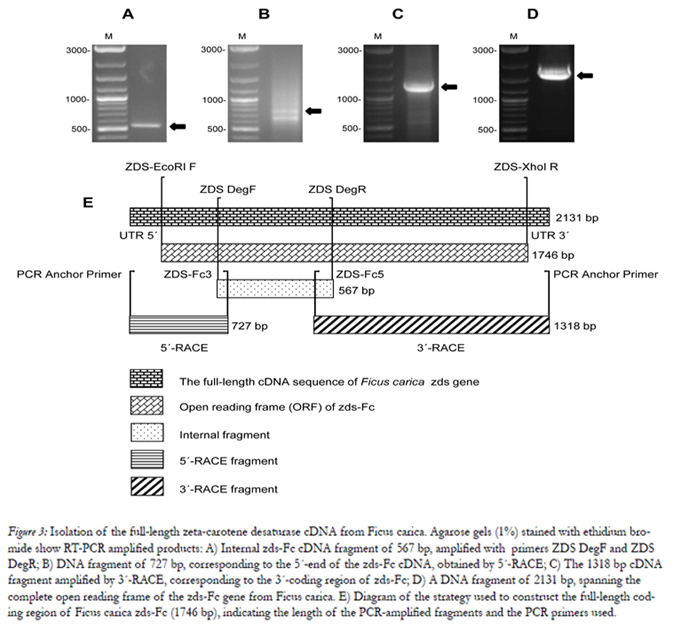
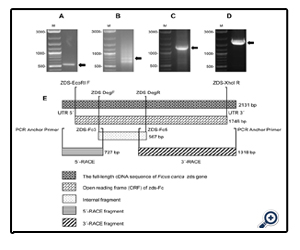
Figure 3: Isolation of the full-length zeta-carotene desaturase cDNA from Ficus carica. Agarose gels (1%) stained with ethidium bromide show RT-PCR amplified products: A) Internal zds-Fc cDNA fragment of 567 bp, amplified with primers ZDS DegF and ZDS DegR; B) DNA fragment of 727 bp, corresponding to the 5´-end of the zds-Fc cDNA, obtained by 5´-RACE; C) The 1318 bp cDNA fragment amplified by 3´-RACE, corresponding to the 3´-coding region of zds-Fc; D) A DNA fragment of 2131 bp, spanning the complete open reading frame of the zds-Fc gene from Ficus carica. E) Diagram of the strategy used to construct the full-length coding region of Ficus carica zds-Fc (1746 bp), indicating the length of the PCR-amplified fragments and the PCR primers used.
Receiver operating characteristic (ROC) curves [plots of sensitivity against (1-specificity)] were plotted to estimate the optimal cutoff value of different values of sensitivity (Se) and specificity (Sp). Area under the ROC curve (AUC) was estimated by two-tailed test with an accuracy value of 0.5 as significance level [19]. The perfect diagnostic test would appear as close as possible to the upper left-hand corner, where both sensitivity and specificity are 100 %. We select a cutoff value of higher sensitivity with minimum false-negative value and high specificity with minimum false-positive [20]. We also measure by the mean optical density of the known negative samples plus two or three times the standard deviation.
The sELISA was evaluated with previously reported commercial ELISA assay (kELISA) and in-house ELISA based on H. pylori surface whole cell antigen (wELISA) [21] by the κ statistic. Student’s t test (one sample and independent samples) was used to compare mean value. Linearity of correlation was calculated by the Pearson’s coefficient correlation method. p < 0.01 were considered as significant. The statistical software, namely SPSS 16.0, MedCalc 11.3.3 and Graph Pad Prism 5.03, was used for analyzing the data.
Tris-HCl buffer SDS-PAGE (Laemmli) was applied to separate the sonicate whole-cell antigen and serum at 10–8 % with 5 % stacking gel by using Bio-Rad Minigel apparatus. Coomassie staining was performed with Coomassie brilliant blue solution [Coomassie brilliant blue (CBB)-R250]. Relative molecular weight (Mr) of unknown protein was determined to calculate their Rf values by using a standard protein marker (Bio-Rad: 161-0318) with linear regression curve method (excel spreadsheet program) [22].
This study was approved by the Karachi University’s Ethical Committee in Karachi, Pakistan.
Results
The strategy used to construct the full-length coding region of Ficus carica zds (zds-Fc) is outlined in Fig.3, and the primers used for this purpose are listed in Table 1. As a first step, we analyzed the published zds sequences, obtained from different plant species, and identified conserved regions in the gene. Based on these regions, we designed degenerate primers ZDS DegF and ZDS DegR. We used these primers to RT-PCR amplify the RNA we isolated from Ficus carica leaves, and this generated a DNA fragment of 567 bp (Fig. 3A). By sequence analyses, the amplified DNA was shown to span to the central area of zds-Fc, and displayed a high similarity (83% to 80%) to other plant zeta-carotene desaturase genes, such as those from Citrus unshi (AB072343.1), Citrus maxima (EU798286.1), Malus x domestica (AF429983.1), Citrus sinensis (AJ319762.1), Jatropha curcas (GQ337075.1), Citrus x paradisi (AF372617.1), Fragaria x ananassa (FJ795343.1), Carica papaya (FJ812088.1) and Daucus carota (DQ192189.1).
The missing 5´and 3´ regions of zds-Fc were then obtained by RACE-PCR, using the Ficus carica cDNA, enerated above by RT, as template, and using primers designed from either the 5’ or 3’ area, respectively, of the above described central area of zds-Fc. This resulted in a 727 bp encompassing the 5’-coding region of the gene (Fig. 3B), and a 1318 bp spanning the missing 3’-coding region of zds-Fc (Fig. 3C). The DNA fragments also share a common area, at either the 5’ or 3’-end, respectively, with the central part of the zds-Fc gene, used to design the PCR primers (Fig.3E). Hence, the three DNA fragments together span the full-length coding region of zds-Fc, and were used to PCR amplify a 2131 bp DNA fragment. Apart from the zds-Fc coding region, the PCR product also contained 197 nucleotides of 5´-UTR and 188-nucleotides of 3´-UTR (data not shown). Finally, we sequenced the zds-Fc cDNA from Ficus carica and deposited it in as the GenBank data base (accession number of JN896309). The complete open reading frame (ORF) of the gene has a length of 1746 bp (Fig. 3D).
The zds-Fc gene coded for a polypeptide of 582 amino acids, with a estimated molecular weight of 64 kD. The ZDS-Fc polypeptide sequence was compared to published zeta-carotene desaturase proteins from other plants. ChloroP 1.1 and TargetP 1.1 predicted a putative chloroplast transit peptide targeting sequence, encompassing the N-terminal amino acids 1 to 43 of ZDS Fc; this correlates well with the transit peptides found in maize, Arabidopsis, Narcissus, and Triticum, which are 30, 34, 42 and 32 residues long, respectively (Al-Babili et al. 1998; Cong et al. 2009; Matthews et al. 2003). The PSORT program predicted a putative polypeptide cleavage site, located: between amino acids 33 and 34, whereas PSIpred found, in our F. carica protein (ZDS Fc), a highly conserved motif characteristic of higher plants ZDS enzymes. Another conserved motif present in ZDS Fc is a typical pyridine dinucleotide binding domain (FAD-binding domain), with a secondary structure of β-sheet-α-helix loop-β-sheet, that is also characteristic of other plant ZDSs (Matthews et al. 2003; Wirenga et al. 1986).
The degree of identity (and homology) between the ZDS-Fc polypeptide and other plant ZDSs was examined by BlastP search. This analysis revealed that our enzyme has an 84% amino acid identity (91% similarity) with Jatropha curcas, 85 % (91%) with Citrus maxima, 85% (91%) with Citrus unshiu, 84% (91%) with Citrus sinensis, 84% (90%) with Malus x domestica, 83% (91%) with Fragaria x ananassa, and 83% (90%) with Carica papaya. These data, taken together, confirm the cloned zds-Fc as an integral part of the ZDS family of proteins.
Expression of Ficus carica zds was confirmed by SDS/ PAGE analyses of E. Coli lysates transformed with the plasmid pET21a, either alone, or containing the complete coding area of the zds-Fc gene (E. Coli BL-zds- Fc, Table 2). A protein with a molecular weight of 64 kD was observed only in the bacterial cells expressing zds-Fc, and only after protein expression was induced by addition of IPTG (data not shown).
We conducted a phylogenetic study of ZDS-Fc on the basis of its predicted polypeptide sequence and that of a variety of clone plant ZDS sequences, available from GeneBank. The phylogenetic tree, shown in Fig. 2, was inferred using the Neighbor-Joining method (Saitou and Nei 1987). This test showed that our expressed protein has a high homology to other plant zeta- carotene desaturases. The zeta-carotene desaturase from marine microalgae Dunaliella salina was included as an out-group and, hence, appears in an isolated branch. The phylogenetic analyses clearly demonstrated that Ficus carica ZDS (JN896309) clusters better with Fragaria x ananassa (FJ795343.1), Malus x domestica (AF429983.1), Carica papaya (FJ812088.1), Jatropha curcas (GQ337075.1), Citrus maxima(EU798286.1), Citrus sinensis (AJ319762.1), and Citrus unshiu (DQ309869.1), than with the other ZDSs included in Fig. 2. Contrary to expectations, the monocot ZDS proteins from Narcissus tazetta var. chinensis and Oncidium Gower Ramsey were not more closely related to the grass carotenoid desaturase than the ZDSs from dicot plants, such as Arabidopsis thaliana or Brassica rapa. The Bootstrap values on the nodes (Fig. 2) indicate the number of times that each group accurred with 1000 replicates. Models with the lowest BIC scores (Bayesian Information Criterion) are considered to describe the substitution pattern as best. So, the evolutionary distances were computed using the Jones-Taylor-Thornton method (Nei and Kumar 2000). The rate variation among sites was modeled with a Gamma distribution (shape parameter = 1) with 5 rate categories, and by assuming that a certain fraction of the sites are evolutionarily invariable.
We constructed a recombinant E. Coli strain (E. Coli BL-pEBI, Table 2), encoding GGPP and phytoene synthases, as well as phytoene desaturase, hence producing lycopene. We used the cell extracts from this bacterial culture, expressing lycopene, as a positive control for our HPLC analyses. We also constructed two recombinant E. Coli strains (E. Coli BL-pEBP and E. Coli BL-pEBR, Table 2) to use as negative controls. Induction of recombinant protein expression in E. Coli BL-pEBP should result in the accumulation of ζ-carotene, which can be used as a substrate by zeta carotene desaturase. On the other hand, the recombinant E. Coli BL-pEBR should produce neurosporene. HPLC analyses confirmed the carotenoids present in the bacterial extracts from our engineered controls as zeta-carotene (E. Coli BL-pEBP), neurosporene (E. Coli BL-pEBR) and lycopene (E. Coli BL-pEBI).
In order to test the enzymatic activity of our recombinant zds-Fc, we constructed two more recombinant E. Coli strains (E. Coli BL-EBPZ and E. Coli BLEBRZ, Table2). The only difference between the negative control E. Coli BL-pEBP and E. Coli BL-EBPZ is the presence in the latter of an additional plasmid (pET21-zdsFc), encoding Ficus carica ZDS. Production of a functional ZDS-Fc enzyme by E. Coli BL-EBPZ would result in the conversion of the zeta-carotene, produced by E. Coli BL-pEBP, into neurosporene and lycopene (Fig. 1). For its part, E. Coli BL-EBRZ is only different from the negative control (E. Coli BL-pEBR) by the presence in the former of an additional plasmid (pET21-zdsFc), encoding Ficus carica ZDS. As above, production of a functional ZDS-Fc enzyme by E. Coli BL-EBRZ would result in the conversion of the carotenoid accumulated by E. Coli BL-pEBR (neurosporene) into the product of ZDS activity, in this case lycopene (Fig. 1). As expected, ZDS expression by E. Coli BL-EBPZ resulted in the enzymatic conversion of part of the ζ-carotene, accumulated by the E. Coli BLEBP culture, into neurosporene and lycopene. This was confirmed by the HPLC detection of two new compounds, not present in the E. Coli BL-pEBP culture. The HPLC elution profile for these compounds recorded maximum absorbance peaks at 416, 441 and 471 nm (identified as neurosporene) and 445, 472, and 504 nm (identified as lycopene). Accordingly, ZDS expression by E. Coli BL-EBRZ resulted in the enzymatic conversion of part of the neurosporene accumulated by the E. Coli BL-EBR culture into lycopene, a compound not present in the E. Coli BL-pEBR culture. The new pigment was confirmed as lycopene by HPLC analysis, and exhibited the expected maximum absorbance peaks at 445, 472 and 504 nm. These results, taken together, confirm the enzymatic ability of ZDS to act on both zeta-carotene and neurosporene, as expected from a plant zeta-carotene desaturase.
The neurosporene content of the recombinant E. Coli BL-EBPZ culture was estimated at 34 μg per g, of cells (dry weight) and the lycopene content at 11 μg per gram of cells (Table 3). Whereas, the lycopene content of the recombinant E. Coli BL-EBRZ culture was estimated at 13 µg per gram of cells (dry weight). Carotenoid production was also apparent by the accumulation of pigment in the recombinant E. Coli cultures and was found to be dependent on the temperature at which the bacteria were grown. Both E. Coli BL-EBPZ and E. Coli BL-EBRZ cultures accumulated most pigment when grown at 28°C in the dark.
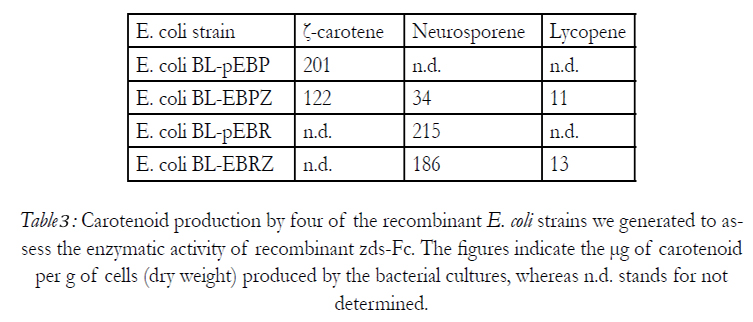
Table 3: Carotenoid production by four of the recombinant E. coli strains we generated to assess the enzymatic activity of recombinant zds-Fc. The figures indicate the μg of carotenoid per g of cells (dry weight) produced by the bacterial cultures, whereas n.d. stands for not determined.
Discussion
Carotenoids are widely distributed natural pigments responsible for the yellow, orange and red colors of fruits, roots and flowers, as they invariably occur in the chloroplasts of higher plants. But the importance of carotenoids in food goes far beyond their role as natural pigments. Carotenoids are not only the starting material for the synthesis of vitamin A, an essential vitamin, but also carry out a variety of biological functions independent of the provitamin A activity (Fraser and Bramley 2004) , attributed to the antioxidant property of carotenoids resulting in deactivation of free radicals (Richmond 2000). They have been reported to enhance the immune system, as well as decrease the risk of degenerative diseases such as cancer, cardiovascular disease, age-related macular degeneration, and cataract formation (Fraser and Bramley 2004; van den Berg et al. 2000).
With consumers these days looking for natural products that would offer them more health benefits, carotenoids emerge as a logical target for the food industry. Plant carotenoid production is insufficient to satisfy this market; hence it is essential to generate a sustainable alternative capable of producing large amounts of natural, high quality carotenoids. Carotenoid production can be rapidly increased by the use of recombinant DNA technology, but before this can be done, the carotenoid biosynthetic genes from many different plants and other organisms need to be cloned and characterized. Moreover, heterologous expression of biosynthetic pathways in E. Coli continues to be a powerful approach for developing metabolic engineering applications in plants. The utility of the bacterial system lies in its inherent similarity to the biochemistry of the plant plastid (Gallagher et al. 2003), thus generating carotenoids that structurally identical to those produced by plants.
Here, we describe the cloning and characterization of a novel ζ-carotene desaturase from the fig tree Ficus carica. Our strategy was to analyze the published zds sequences, from other plant species, identify conserved regions in the gene and design degenerate PCR primer that were used to clone the central region of the zds- Fc gene. The rest of the gene was then obtained by RACE-PCR. The sequence amplified for the zeta-carotene desaturase gene from Ficus carica is 2131 bp long and spans a 1746 bp open reading frame, encoding a protein of 582 amino acid residues, with an estimated molecular weight of 64 kD.
The ZDS-Fc polypeptide sequence was compared to published zeta-carotene desaturase proteins from other plants and was found to share many characteristics common to other plant ZDS s so, as it is the case for other enzymes in the carotenoid biosynthesis pathway, ZDS appears to be highly conserved in higher plants (Cong et al. 2009). Another common characteristic was the presence in ZDS of an N-terminal putative chloroplast transit peptide targeting sequence, which was predicated to be 43 residues long, this is also typical of other enzymes involved in carotenoid biosynthesis in plants (Bouvier et al. 2005; Fraser and Bramley 2004; Sandmann et al. 2006) . Finally, the sequences also contained a typical pyridine dinucleotide binding domain (FAD-binding domain), with a secondary structure of β-sheet-α-helix loop-β-sheet, that is characteristic of other plant ZDSs (Matthews et al. 2003; Wirenga et al. 1986). These data, taken together, confirms the cloned zds-Fc as an integral part of the ZDS family of proteins.
Based on the shared sequences between the F. carica enzyme and other plant ZDSs, we constructed a phylogenetic tree illustrating the evolutionary relationship between the fig tree ZDS and other published carotenoid zeta carotene desaturases from higher plants. Ficus carica ζ-carotene desaturase was further characterized as part of the ZDS family of plant enzymes, and it clustered better with the dicot plant trees ZDSs such as Citrus unshiu, Jatropha curcas, Carica papaya and Malus x domestica.
Recombinant expression of Ficus carica zds in E. Coli produced a protein with a molecular weight of 64 kD, consistent with that estimated from the amino acid sequence of the polypeptide. In order to test the in vivo enzymatic activity of our recombinant zds-Fc, we engineered four recombinant E. Coli strains (E. Coli BLEBP BL-EBR, BL-EBPZ, and BL-EBRZ, Table2). Whereas the first two bacterial cultures only accumulated ζ-carotene and neurosporene, respectively, the last two recombinant strains (E. Coli BL-EBPZ, and BL-EBRZ), expressing our recombinant zds-Fc, were able to convert the ζ-carotene and neurosporene into neurosporene and lycopene, or lycopene, respectively. These results demonstrated, in vivo, that ZDS, a single desaturase plant enzyme, efficiently catalyzed the last two desaturation steps leading to the production of Lycopene (Table 3). Hence, as described for other plant enzymes (Al-Babili et al. 1998; Cong et al. 2009; Matthews et al. 2003; Yan et al. 1986), the Ficus carica zds gene we cloned encodes a zeta-carotene desaturase which is capable of catalyzing the conversion of ζ-carotene into first neurosporene and then lycopene. In summary, this study, represents the first time that the Ficus carica ζ-carotene was, not only cloned and characterized, but was also shown, by recombinant expression in E. Coli, to catalyze the desaturation of ζ-carotene to produce first neurosporene and finally lycopene.
The authors wish to thank Dr. Norihiko Misawa (Research Institute for Bioresources and Biotechnology, Ishikawa Prefectural University, Japan) for the gift of plasmids pACCRT-EBR and pACCRT-EBI; and Dr. Gerhard Sandmann (J. W. Goethe-Universität, Frankfurt, Germany) for the gift of plasmid pACCRT-EBP. Thanks are also due to both the Faculty of Pharmacy and the School of Biotechnology for their support throughout this project.
References
- Al-Babili S, Oelschlegel J, Beyer P (1998) Plant Gene Register PGR98-103: A cDNA encoding for zeta-carotene desaturase from Narcissus pseudoNarcissus L. Plant Physiol 117:717–720
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic Local Alignment Search Tool. J.Mol.Biol 215(3):403- 410
- Araya-Garay JM, Feijoo-Siota L, Veiga-Crespo P, Villa TG (2011) cDNA cloning of a novel gene codifying for the enzyme lycopene β-cyclase from Ficus carica and its expression in the Escherichia coli. Appl Microbiol Biotechnol 92:769-777
- Bouvier F, Rahier A, Camara B (2005) Biogenesis, molecular regulation and function of plant isoprenoids. Prog.Lipid Res 44(6):357-429
- Cong L, Wang C, Li Z, Chen L, Yang G, Wang Y, He G (2009) cDNA cloning and expression analysis of wheat (Triticum aestivum L.) phytoene and zeta-carotene desaturase genes. Mol Biol Rep. Doi:10.1007/s11033-009-9922-7
- DellaPenna D, Pogson BJ (2006) Vitamin synthesis in plants: Tocopherols and carotenoids. Annu Rev Plant Biol57(1):711-738fg
- Emanuelsson O, Henrik N, Soren B, Gunnar von Heijne (2000) Predicting subcellular localization of proteins based on their N-terminal amino acid sequence J Mol Biol 300:1005-1016
- Emanuelsson O, Nielsen H, Von Heijne G (1999) ChloroP,a neural network-based method for predicting chloroplast transit peptides and their cleavage sites. Protein Sci 8(5):978- 984
- Fraser PD, Bramley P (2004) The biosynthesis and nutritional uses of carotenoids. Prog.Lipid Res 43(3):228-265
- Gallagher CE, Cervantes M, Wurtzel ET (2003) Surrogate biochemistry: use of Escherichia coli to identify plant cDNAs that impact metabolic engineering of carotenoid accumulation. Appl Microbiol Biotechnol 60(6): 713-719
- Hirschberg J (2001) Carotenoid biosynthesis in flowering plants. Curr Opin Plant Biol 4(3):210-218
- Jones DT (1999) Protein secondary structure prediction based on position-specific scoring matrices. J.Mol.Biol 292(2):195-202
- Krinsky N (1989) Antioxidant functions of carotenoids. Free Radical Biol Med 7:617-635
- Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature (London, U. K.) 227:680-685
- Linden H, Vioque A, Sandmann G (1993) Isolation of a carotenoid biosynthesis gene coding for zeta-carotene desaturase from Anabaena PCC 7120 by heterologous complementation. FEMS Microbiol Lett 106:99-104.
- Matthews PD, Luo RB, Wurtzel ET (2003) Maize phytoene desaturase and f-carotene desaturase catalyse a poly-Z desaturase pathway: implications for genetics engineering of carotenoid content among cereal crops. J Exp Bot 54:2215– 2230
- Melendez Martinez AJ, Vicario IM, Heredia FJ (2003) A routine high-perfomance liquid chromatography method for carotenoid determination in ultrafrozen orange juices. J Agric Food Chem 51:4219-4224
- Misawa N, Shimada H (1998) Metabolic engineering for the production of carotenoids in non-carotenogenic bacteria and yeasts. Journal of Biotechnology 59(3):169-181
- Misawa N, Nakagawa M, Kobayashi K, Yamano S, Izawa Y, Nakamura K, Harashima K (1990) Elucidation of the Enwinia uredovora carotenoid biosynthetic pathway by functional analysis of gene products expressed in Escherichia coli. J Bacteriol 172:6704-6712
- Nakai K, Horton P (1999) PSORT: A program for detecting sorting signals in proteins and predicting their subcellular localization. Trends Biochem Sci 24(1): 34-35
- Nei M, Kumar S (2000) Molecular Evolution and Phylogenetics. Oxford University Press, New York.
- Nielsen H, Engelbrecht J, Brunak S, von Heijne G (1997) Identification of prokaryotic and eukaryotic signal peptides and prediction of their cleavage sites. Protein Engineering 10:1-6
- Park H, Kreunen SS, Cuttriss AJ, Dellapenna D, and Pogson BJ (2002) Identification of the carotenoid isomerase provides insight into carotenoid biosynthesis, prolamellar body formation and photomorphogenesis. Plant Cell 14:321-332
- Peelman F, Labeur C, Vanloo B, Roosbeek S, Devaud C, Duverger N, Denèfle P, Rosier M, Vandekerckhove J, Rosseneu M (2003) Characterization of the ABCA transporter subfamily: identification of prokaryotic and eukaryotic members, phylogeny and topology. J Mol Biol 325(2):259-274
- Richmond A (2000) Microalgal biotechnology at the turn of the millennium: A personal view. J Appl Phycol 12:441-451
- Rodrigo MJ, Marcos JF, Zacarías L (2004) Biochemical and molecular analysis of carotenoid biosynthesis in flavedo of orange (Citrus sinensis L.) during fruit development and maturation. J Agric Food Chem 52(22):6724-6731
- Rodríguez Concepción M (2006) Early steps in isoprenoid biosynthesis: Multilevel regulation of the supply of common precursors in plant cells. Phytochemistry Reviews 5(1):1-15
- Saitou N, and Nei M (1987) The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol 4:406-425
- Sandmann G, Romer S, Fraser PD (2006) Understanding carotenoid metabolism as a necessity for genetic engineering of crop plants. Metab Eng 8(4):291-302
- Sandmann G (2002) Molecular evolution of carotenoid biosynthesis from bacteria to plants. Physiol Plant 116:431-440
- Sandmann G (1994) Phytoene desaturase - Genes, enzymes and phylogenetic aspects. J Plant Physiol 143(4-5): 444-447
- Takaichi S, Sandmann G, Schnurr G, Satomi Y, Suzuki A, Misawa N (1996) The carotenoid 7,8-dihydro-ψ end group can be cyclized by lycopene cyclases from the bacterium Erwinia uredovora and the higher plant Capsicum annuum. Eur J Biochem 241:291-296
- Tamura K, Peterson D, Peterson N, Stecher G, Nei M, and Kumar S (2011) MEGA5: Molecular Evolutionary Genetics Analysis using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Molecular Biology and Evolution 28:2731-2739
- van den Berg H, Faulks R, Granado HF, Hirschberg J, Olmedilla B, Sandmann G, Southon S, Stahl W (2000) The potential for the improvement of carotenoid levels in food and the likely systemic effects. J Sci Food Agric 80(7):880-912
- Wirenga RK, Terpstra P, Hol WGJ (1986) Prediction of the occurrence of the ADP-binding β´β-fold in proteins, using an amino acid sequence fingerprint. J Mol Biol 187:101-107
- Yan P, Gao XZ, Shen WT, Zhou P (2011) Cloning and expression analysis of phytoene desaturase and ζ-carotene desaturase genes in Carica papaya. Mol Biol Rep 38:785-791.





