Exploring Bilateral Asymmetry & Ante-Mortem Trauma in Pleistocene Narmada Hominin Fossil Clavicles
Dr. Anek Ram Sankhyan1, 2*
1 Anthropologist (Physical) & Visiting Fellow, Anthropological Survey of India, Kolkata (rtd.), India.
2 PresentlyatPalaeo Research Society, Ghumarwin-174021 (H.P.), India.
*Corresponding Author
Dr. Anek Ram Sankhyan,
Anthropologist (Physical) & Visiting Fellow, Anthropological Survey of India, Kolkata (rtd.), India.
PresentlyatPalaeo Research Society, Ghumarwin-174021 (H.P.), India.
E-mail: arsankhyan@gmail.com
Received: May 27, 2022; Accepted: June 21, 2022; Published: June 28, 2022
Citation: Dr. Anek Ram Sankhyan. Exploring Bilateral Asymmetry & Ante-Mortem Trauma in Pleistocene Narmada Hominin Fossil Clavicles. Int J Forensic Sci Pathol. 2022;9(4):495-499.
Copyright: Dr. Anek Ram Sankhyan�2022. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
The present study explores bilateral asymmetry accountable to ante-mortem trauma in two mid-late Pleistocene hominin clavicles discovered from Hathnora in the central Narmada valley, which had earlier yielded a partial cranium. The two female fossilized clavicles and the female cranium were initially thought of belonging to the same individual, but of late, it was confirmed that they belong to two different hominins. The clavicles come from a pygmoid/ short stocky female showing intriguing bilateral differences. The 3-D scans and comparative metric and non-metric analyses reveal that the two clavicles exhibit considerable bilateral asymmetry accountable for the left shoulder ante-mortem trauma and some years of post-trauma survival to allow healing of the broken end.
2.Introduction
3.Observations
4.Discussion And Conclusions
5.Acknowledgements
6.References
Introduction
Several experts have different interpretations of the functional
anatomy and peculiarities of the human clavicles [9, 8, 16, 15,
27, 19, 28, 2]. The most common interpretation is that it functions
as a strut bone connecting the upper limb with the axial
skeleton transferring the upper-limb forces to the thorax. But,
at the same time the clavicle stabilizes the gleno-humeral joint
by limiting motion of the scapula and anchoring the joint to the
ribs and sternum through the sternoclavicular joint. This happens
with the help of six muscles, such as pectoralis major, deltoid,
sternocleidomastoid, sternohyoid, trapezius, and subclavius, and
four ligaments which support as well as transmit forces and also
give it sigmoid or S-shaped or two smooth curves which may vary
in depth. Thus, the clavicle endures a variety of loads, torsion
and shear stresses and modeled to compressive (axial) and bending
loads, resulting from weight and compression exerted by the
upper limb on the glenoid. Variation in the cross-sectional shape
along the bone reflects this variety of mechanical loadings supporting
the multiple functional capacities of the clavicles.
Human clavicle is also unique in its development compared to
other postcranial bones. It is the only postcranial bone to develop,
in part, from membranous (i.e., ��dermal�� cell layer) tissues,
as opposed to chondral models [39, 27]. It is also the first bone
to ossify in the upper-limb complex and attains early much of
its adult morphology during foetal growth, and its �S� or doublecurved
shape is achieved well before birth, leaving further growth
to occur primarily at the ends, especially the medial epiphysis [34].
Having much developed in utero, the clavicle is among the last
bones to reach complete ossification of the epiphyses [33]. It has
a developmental pathway that is different from the humerus, and
therefore has a much longer period to acquire life style changes
and adaptations to lateralized behaviours (bilateral asymmetry)
relative to the humerus and radius.The clavicle has a great range
of variation in size, in the development of the conoid and deltoid
tubercles, the epiphyses as well as the rhomboid fossa. Modern
human clavicle is associated with a higher scapula in relation to
the thorax, while it is, lower in position in the Neanderthal. There
are three main types of shapes in dorsal view [31, 32].
Observations
The author [20, 21, 23] reported two mid-late Pleistocene hominin
clavicles from Hathnora in the Central Narmada valley, which had
earlier yielded a partial female cranium [37]. The two fossilized
clavicles were also sexed as belonging to a female and were therefore
initially thought of belonging to the same individual [22], but of late, through excavations at Hathnora, it was confirmed that
they come from two different stratigraphic and cultural contexts,
and belong to two different hominins [6, 24-26].
The right clavicle
The right clavicle was initially confused to its sternal epiphysis
partly broken off [20] owing to the narrow sternal end contrasting
with modern humans, and therefore its maximum length was
estimated to be 100 mm. But, soon it is found to be remarkably
complete, well-preserved and extremely rare fossil of its kind with
90 mm maximum length, confirmed through the 3D Scans (Fig.
1) revealing that the sternal articular capsule and the facets are
indeed intact.Its diaphysis is slightly flattish with S-shaped or sigmoid
curves typical of a human clavicle, although itsshortness
and shallower curves give the clavicle a straighter look with little
axial torsion comparable to the Andaman Pygmy [20] (Fig. 1C).
The M. pectoralis major attachment on the anterosuperior aspect
for the proximal diaphysis is rugose and distinct.The posterior
diaphyseal border is rough and undulating but sharpens toward
the eroded proximal part. The M. subclavius attachment area is
distinctly rugose and undulating. The M. sternocleidomastoideus
area located posterosuperiorly on the proximal diaphyseal end is
distinct and relatively flat. On the posteroinferior aspect of the
proximal diaphysis, M. sternohyoideus attachment is also well developed.
The costoclavicular ligament facet (rhomboid fossa) is
located centrally on the inferior surface of the proximal diaphysis
because of the flatness and little no axial torsion of the bone. Due
to some erosion it extends almost to the midpoint of the bone
is quite developed (21.9 x 8.0 mm) furrow and bilipped suggesting
strong muscular attachment of the clavicle with the rib. The
subclavian groove is like a deep and broad sulcus in the concavity
of the inferior surface of the distal epiphysis and is anteriorly
disposed. However, it becomes shallow and indistinct when it
reaches the middle of the diaphysis.


Figure.1. 3-D Scans of the Narmada Right Clavicle (complete) (A) and Left Clavicle (partial) (B) in four views,
C. Scans of the Andaman Pygmy right and left clavicles
D: Metric Landmarks on Right Clavicle: 1-8 are various antero-posterior diameters in caudal plane; AB= maximum length, CD=Conoid length.
The left clavicle
It preserves only the medial 2/3rd portion and lacks the lateral 1/3rd, i.e., the acromial epiphysis. The latter is broken off beyond the conoid tubercle, which is the common weak point of the clavicle. The broken end is smoothly rounded and blunt as if healed from an ante mortem trauma suffered during the lifetime of the hominin. The medial end is well preserved and possesses the sternal articular capsule and the facet typical of the clavicle. It is completely mineralized as the right clavicle and shares the same ash-gray hue, also shared by the rib fossil, all recovered from the same site.
Age and Sex of the clavicles
Both the clavicles are quite thick unlike the thinner juvenile modern clavicles and quite apparent of being of adult physiological age as revealed by the developed muscular rugosities. The sternal epiphysis-diaphysis fusion detection is difficult in the fossilized bones as the X-rays cast complete continuous shadow as if completely fused; the complete diaphyseo-epiphyseal fusion in modern human clavicles is generally observed having occurred between 25 -35 years of age [13, 10, 11, 7, 14, 33]. So, if modern standards hold true, the Narmada clavicles may belong to an adult individual of above 25 years of age at death.
Probably modern non-metric sexing criteria may equally hold well for the Middle Pleistocene Narmada Homo because the early Pleistocene African Homo erectus (=H. ergaster) shows modern upper chest morphology as observed by [8]. The Narmada clavicles also show advance trend of moderate rounding of the shafts (mesocleidy) approaching the lower threshold of Homo sapiens. Therefore, we may apply the modern sexing criteria to know the sex of Narmada clavicles. On these criteria both the Narmada clavicles may be sexed as of female as they show: (i) the narrow sternal end or a lesser Inner-End Index -a sum of the antero-posterior and supero-inferior diameters at the sternal end [17]; (ii) the conspicuous conoid tubercle; (iii) the smaller acromial facet- quite evident on the otherwise robust / male-like looking right clavicle; (iv) the straighter or less curved diaphysis of both clavicles; (v) the unusually short clavicle length [35, 38, 36].
But, the metrical criteria to differentiate the two sexes as suggested by [13, 11], for various north Indian populations are inapplicable for the very short clavicles. However, the clavicles of the female pygmies of the shortest Andaman Islanders, namely the Onge of the Little Andaman and the Greater Andamanese of Middle Andaman, show similar length (Table 1) and also approach in general morphology, especially in the lesser curvature and the rounded deltoid region exhibited by the left Narmada clavicle.
Comparative analysis of the clavicles
The Narmada clavicles were compared with a mixed-sex sample of 30 adult modern clavicles (14 modern mainland humans and 16 Andaman Islanders) besides incorporating a few ancient Indian clavicles in the palaeoanthropological repository of the Anthropological Survey of India, Kolkata. The pygmy sample is from the Onge- the forest dweller Negrito forager of the Andaman Islands, who now number less than a 100. The clavicles of the modern human sample represent a heterogeneous eastern Indian population in the repository of the Anthropological Survey of India, Kolkata. In all the clavicles were studied for 36 mensural traits of which five constitute indices, besides for five non-mensural traits (Table 2). The selection and methods of measurements of the variables were as per the criteria followed by [4, 30, 3, 18, 17], besides more additions in the variables.
Bilateral differences and Asymmetry
Compared to the right clavicle, the left clavicle presents some of its own uniqueness, which could easily detract one to assign it to the same individual as represented by the right clavicle. These are: (i) the diaphysis is more rounded and gracile with more axial twist; (ii) the sternal epiphysis and the articular facet more rounded; (iii) as a consequence of the above, the conoid tubercle is conspicuous, the deltoid ridge and deltoid tuberclerounded,and fine narrow subclavian groove; (iv) on the contrary, the right clavicle has a sharp deltoid ridge leading to diminished deltoid tubercle, and possesses a deep subclavian and costoclavicular attachment facet (rhomboid fossa), which in the left clavicle is less distinct and infero-laterally disposed unlike the centrally located one of the right clavicle; (v) the muscular rugosities of the M. sternocleidomastoideus, M. subclavius and M. pectoralis majorare also more developed in the right clavicle than in the left; (vi) the anteromedial border of the left clavicle is slightly uneven compared to that of the right clavicle.
The great distinction between the two fossil clavicles was initially argued to two different individuals. But, it was unlikely that two females are represented in the same site,one leaving the right and the other the left clavicle. But, why so much bilateral difference, prompted comparative analysis presented in Table 1 and Table 2.
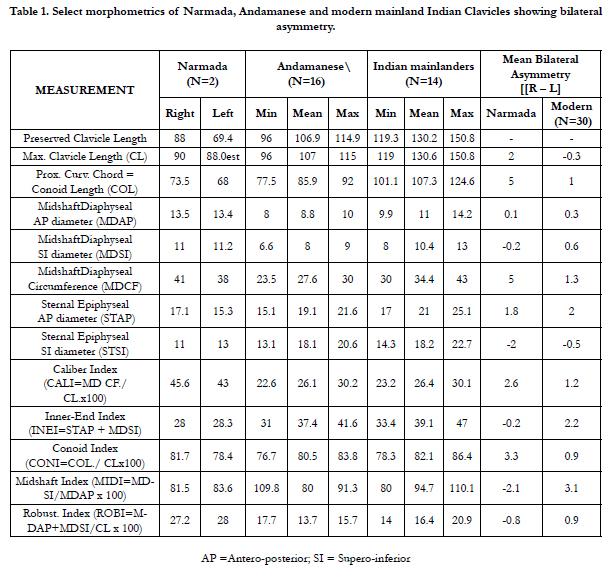

Table 1. Select morphometrics of Narmada, Andamanese and modern mainland Indian Clavicles showing bilateral asymmetry
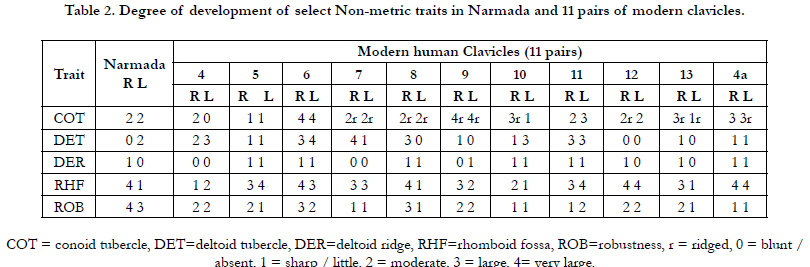
Table 2. Degree of development of select Non-metric traits in Narmada and 11 pairs of modern clavicles.
Discussion And Conclusions
The main points in favour of the two clavicles coming from the
same individual are: the similar inferred adult age, the same female
sex, and the same siteand similar gray hue, similar maximum clavicular
lengths (90 mm for right clavicle and 88 mm for the left clavicle) also suggesting contemporaneity. In addition, the degree
of robusticity is also similar in the two clavicles as revealed from
their robusticity and caliber indices and by other mensural traits
shown in Table 1, which also incorporates the data for 30 modern
clavicles. The Table 2 reveals considerable non-metric bilateral
variation in modern clavicles for various parameters, which is
noteworthy in the development of the rhomboid fossa, the deltoid
and conoid tubercles, and the deltoid ridge. The two Narmada
clavicles also exhibit considerable bilateral variation, and therefore,
the likelihood of occurrence of one left and another right
clavicle of two different adult females at the same siteis ruled out.
Human clavicular asymmetry is significantly left-biased in length
and right-biased in diaphyseal breadth [2, 1, 16, 4, 6]. This length
asymmetry is contra lateral to the length asymmetry observed
in humeri in the same sample, though clavicles and humeri have
same-side directional asymmetry in diaphyseal breadths. This pattern
provides more support for the notion that the clavicle is an
integral functional element of the upper-limb complex. As in previous
research of all the major long bones of the postcranium,
the diaphyseal breadths were found to exhibit greater amounts of
asymmetry than the long-bone lengths. Despite some variation,
these patterns are universal among all modern humans sampled.
Females have significantly greater directional asymmetry in humeral
length, whereas males have significantly more directional
humeral diaphyseal asymmetry, but this sexual dimorphism is not
evident in the clavicle. Results further suggest that different regions
of the bones react differently to behaviour (i.e., mechanical
loading) and/or genetic influences. The clavicle, although a
functional component of the upper limb, has a developmental
pathway that is different from the humerus, which therefore results
in some discontinuity in shared patterns of asymmetry between
these bones. Diaphyseal breadths however, continue to
exhibit greater sensitivity to loading than lengths of long bones
[2]. Comparisons of groups that had different patterns of activity
and loading behaviors emphasize the observable effects of these
differences among patterns of asymmetry.
There are many studies, which support presence of bilateral variation
in the clavicle due to a differential use of the two shoulders/
arms employed habitually in various occupations or life styles,
which result in more variation than the climate [18]. For example,
a distinctly deep and prominent �rhomboid fossa� is regarded a
clear sign of �handedness� [10, 29]. In view of this, the deeper
rhomboid fossa on Narmada right clavicle suggests a �distinctly
right-handed� individual- who likely �over-used� the right shoulder/
arm.
There are additional morphological traits preserved on Narmada
clavicles which further attest bilateral asymmetry as follows: (i)
shelf-like ex-ostosis at the M. sternocleidomastoideus at the medial
curvature, which adds to its strength; (ii) Rugged M. subclavius
and deep subclavian groove- having turned into a sulcus; (iii)
expanded and sharp deltoid ridge lacking the deltoid tubercle; (iv)
strongly developed anterior diaphyseal border having turned into
a tiny keel or a linear mid-rib.
The above features would support very strong shoulder and chest
muscles of the right side. These peculiar developments probably
indicate stress-induced anomalous bilateral asymmetry with the
left side less in use. This may also suggest that the hyper- robustness
of the Narmada clavicle could be an individual peculiarity
attributed to unusual stressful bilateral asymmetry, though the robustness
in general may also be a characteristic of the �archaic�
hominins and of the �short and stocky� or �Pygmoid� hominins.
It appears quite likely that the bilateral asymmetry could have resulted
from the injured left clavicle, i.e., theante mortem traumawhich
consequently led to an �over-use� of the right side. The
broken lithified end of the left clavicle attests a pre-fossilization
fracture of the left clavicle/shoulder. The mineralized distal end is
smoothly rounded or blunt as generally seen in healed up bones,
suggesting therefore, that the hominin probably lived for quite
some time with the injured left shoulder before death.
Acknowledgements
The author conducted the study in the Palaeoanthropology Laboratory
of the Anthropological Survey of India, Head Office at
Kolkata, and thanks Dr. V.R. Rao, the then Director-in-charge, for
the logistic support. He also thanks Professor InduTalwar as the
present work is a part of his PhD thesis submitted to the Panjab
University Chandigarh.
References
- [1]. Auerbach BM, Ruff CB. Limb bone bilateral asymmetry: variability and commonality among modern humans. J Hum Evol. 2006 Feb;50(2):203- 18. PubMed PMID: 16310833.
- Auerbach BM, Raxter MH. Patterns of clavicular bilateral asymmetry in relation to the humerus: variation among humans. J Hum Evol. 2008 May;54(5):663-74. PubMed PMID: 18035397.
- Carretero JM, Arsuaga JL, Lorenzo C. Clavicles, scapulae and humerifromthe Sima de los Huesos site (Sierra de Atapuerca, Spain). J HumEvol. 1997 Aug-Sep;33(2-3):357-408. PubMedPMID: 9300347.
- Churchill SE. Human upper body evolution in the Eurasian Later Pleistocene. The University of New Mexico; 1994.
- Churchill SE, Formicola V. A case of marked bilateral asymmetry in the upper limbs of an Upper Palaeolithic male from Barma Grande (Liguria), Italy. Int J Osteoarchaeol. 1997 Jan;7(1):18-38.
- Churchill SE. Endocrine models of skeletal robusticity and the origins of gracility. Human origins, genome and people of India. 2007 May 16:337- 68.
- Galstaun G. A study of ossification as observed in Indian subjects. Indian J Med Res. 1937;26:267-324.
- Harrington MA Jr, Keller TS, Seiler JG 3rd, Weikert DR, Moeljanto E, Schwartz HS. Geometric properties and the predicted mechanical behavior of adult human clavicles. J Biomech. 1993 Apr-May;26(4-5):417-26. Pub- Med PMID: 8478346.
- Inman VT, Saunders JB. Observations on the function of the clavicle. Calif Med. 1946 Oct;65(4):158-66. PubMed PMID: 18731101.
- Jit I, Kulkarni M. Times of appearance and fusion of epiphysis at the medial end of the clavicle. Indian J Med Res. 1976 May;64(5):773-82. PubMed PMID: 184039.
- Jit I, Sahni D. Sexing the north Indian clavicles. J AnatSoc India. 1983;32(2):61-72.
- Inder J, Singh S. Estimation of stature from clavicles. Indian J Med Res. 1956 Jan;44(1):137-55. PubMed PMID: 13306315.
- Jit I, Singh S. The sexing of the adult clavicles. Indian J Med Res. 1966 Jun;54(6):551-71. PubMed PMID: 5947020.
- Johnston FE. Assessment of growth and age in the immature skeleton. Reconstruction of Life from the Skeleton. 1989.
- Ljunggren AE. Clavicular function. ActaOrthop Scand. 1979 Jun;50(3):261- 8. PubMed PMID: 474097.
- Mays S, Steele J, Ford M. Directional asymmetry in the human clavicle. Int J Osteoarchaeol. 1999 Jan;9(1):18-28.
- Parsons FG. On the Proportions and Characteristics of the Modern English Clavicle. J Anat. 1916 Oct;51(Pt 1):71-93. PubMed PMID: 17103806.
- Pearson OM. Activity, climate, and postcranial robusticity: implications for modern human origins and scenarios of adaptive change. CurrAnthropol. 2000 Aug-Oct;41(4):569-607. PubMed PMID: 11624671.
- Renfree KJ, Wright TW. Anatomy and biomechanics of the acromioclavicular and sternoclavicular joints. Clin Sports Med. 2003 Apr;22(2):219-37. PubMed PMID: 12825527.
- Sankhyan AR. Fossil clavicle of a middle Pleistocene hominid from the Central Narmada Valley, India. J Hum Evol. 1997 Jan;32(1):3-16. PubMed PMID: 9034953.
- Sankhyan AR. A new human fossil find from the Central Narmada basin and its chronology. Current Sci. 1997;73(12):1110-1.
- Sankhyan AR. The place of Narmada hominin in the Jigsaw puzzle of human origins. Gondwana Geological Magazine. 1999;4:335-45.
- Sankhyan AR. New fossils of early Stone Age man from central Narmada valley. Current Sci. 2005 Mar 10;88(5):704-7.
- Sankhyan AR. Pleistocene Hominins & Associated Findings from Central Narmada Valley bearing on the Evolution of Man in South Asia. Ph.D. Dissertation, Chandigarh: Panjab University; 2010.
- Sankhyan AR, Rao VR. Did ancestors of the pygmy or hobbit ever live in Indian heartland. Recent advances on Southeast Asian paleoanthropology and archeology. 2007:76-89. /
- Sankhyan AR. Hominin fossil remains from the Narmada Valley. A Companion to South Asia in the Past. 2016 Jun 8:72-85.
- Cunningham C, Scheuer L, Black S. Developmental juvenile osteology. Academic press; 2016 Jul 26.
- Sellards R. Anatomy and biomechanics of the acromioclavicular joint. OperTechnSportMed. 2004 Jan 1;12(1):2-5.
- Shauffer IA, Collins WV. The deep clavicular rhomboid fossa. Clinical significance and incidence in 10,000 routine chest photofluorograms. JAMA. 1966 Feb 28;195(9):778-9. PubMed PMID: 5951885.
- Vandermeersch B, Trinkaus E. The postcranial remains of the R�gourdou 1 Neandertal: the shoulder and arm remains. J Human Evol. 1995 May 1;28(5):439-76.
- Voisin JL. Clavicle, a neglected bone: morphology and relation to arm movements and shoulder architecture in primates. Anat Rec A DiscovMol Cell Evol Biol. 2006 Sep;288(9):944-53. PubMed PMID: 16894572.
- Voisin JL. Krapina and other Neanderthal clavicles: A peculiar morphology?. PeriodicumBiologorum. 2006 Oct 31;108(3):331-9.
- Webb PA, Suchey JM. Epiphyseal union of the anterior iliac crest and medial clavicle in a ReferencedDec;68(4):457-66. PubMed PMID: 4083337.
- Corrigan G.E. The neonatal clavicle. Biol. Neonatorum 1960; 2: 79-92.
- Dwight T. The Range and Significance of Variation in the Human Skeleton: The Shattuck Lecture for 1894. The Boston Medical and Surgical Journal. 1894 Jul 26;131(4):73-6.
- Schultz AH. Proportions, variability and asymmetries of the long bones of the limbs and the clavicles in man and apes. Human Biology. 1937 Sep 1;9(3):281-328.
- Sonakia A. The skull cap of early man and associated mammalian fauna from Narmada Valley alluvium, Hoshangabad area, Madhya Pradesh, India. Records of the Geological survey of India. 1984;113(6):159-72.
- Terry RJ. The clavicle of the American Negro. American Journal of Physical Anthropology. 1932 Jan;16(3):351-79.
- Wirth MA, Rockwood CA. Acute and chronic traumatic injuries of the sternoclavicular joint. JAAOS-Journal of the American Academy of Orthopaedic Surgeons. 1996 Sep 1;4(5):268-78.
Indexed in





Total Visitors
Copyright © 2019 SciDoc Publishers. All Rights Reserved.
