SIRT7: from a Metabolic Regulator to a Disease Marker
Lin Z
School of Life Sciences, Chongqing University, Chongqing 400044, People’s Republic of China.
*Corresponding Author
Zhenghong Lin Ph.D,
School of Life Sciences,
Chongqing University, Chongqing 400044,
People’s Republic of China.
Article Type: Review Article
E-mail: zhenghong-lin@cqu.edu.cn
Received: February 01, 2015; Accepted: February 19, 2015; Published: February 23, 2015
Citation: Lin Z (2015) SIRT7: from a Metabolic Regulator to a Disease Marker. Int J Cancer Stud Res. 4(1), 63-66. doi: dx.doi.org/10.19070/2167-9118-150009.
Copyright: Lin Z© 2015. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
Acetylation regulation plays an important role in various kinds of physiological processes, dysregulation of which leads to pathological progression and serious diseases. SIRTuin family, a class III HDACs, has drawn more and more attention due to its increasing vital biological functions. In case of cancer progression, some of which has been demonstrated to serve as either tumor suppressor or oncogene or both, which depends on context and/or cell type. Here we summarized the roles of SIRT7 in cancer progression and discussed its therapeutic potential in cancer treatment.
2.Introduction
3.SIRT7 as a Metabolic Modulator
4.SIRT7 as a Deacetylase: Histone and non-Histone Substrates
5.The Role of SIRT7 in Tumorigenesis
5.1.SIRT7 as a Tumor Suppressor?
5.2.SIRT7 as an Oncogene
6.Concluding Remarks
7.Acknowledgement
8.Declaration of Conflicting Interests
8.References
Keywords
SIRT7, Acetylation, Deacetylation, Cancer
Introduction
Acetylation plays an important role in different types of physiological processes including aging, metabolism, apoptosis and neurogenesis, and has been linked to many pathological processes such as cancer, inflammatory and autoimmune diseases. In mammals, acetylation is regulated by histone acetyltransferases (HATs) and histone deacetylases (HDACs). There are four classes of HDACs. Class I HDACs (HDAC1, 2, 3, and 8) consist mainly of a catalytic domain and are widely expressed in mammalian tissues. Class II HDACs are divided into two subclasses, IIa (HDAC4, 5, 7, and 9) and IIb (HDAC6 and 10). Class IV currently contains only HDAC11. SIRTuins belong to the class III histone deacetylases and are nicotinamide adenine dinucleotide (NAD+)- dependent [1]. Mammals have seven SIRTuin members (SIRT1- SIRT7), with the function of SIRT1 being mostly studied in the past decade [2] and others less well studied but attracted more and more attention recently.
All the SIRTuin proteins share a highly conserved NAD+-binding and catalytic core domain, and have deacetylase activity [3], but their subcellular localizations are quite different ([4], table 1). SIRT1, SIRT6, and SIRT7 are predominantly nuclear proteins ([5,6], table 1). SIRT1 can also shuttle between cytosol and nuclear in various tissues [7]. SIRT2 is mainly a cytoplasmic protein while SIRT3, SIRT4, and SIRT5, are primarily found in mitochondria and can regulate mitochondrial activity [8-10].
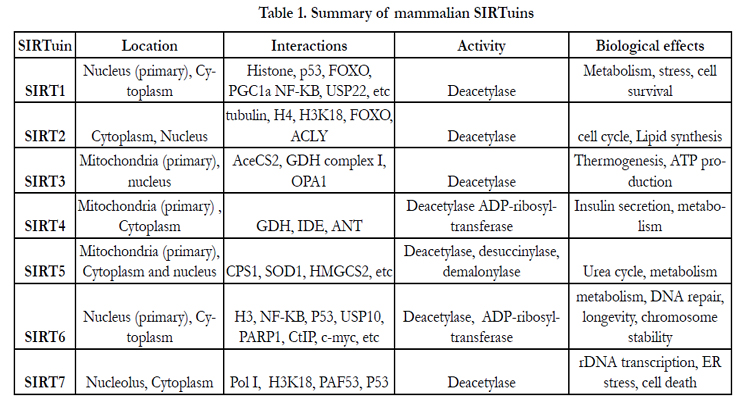

Table 1. Summary of mammalian SIRTuins
In the SIRTuin family, SIRT1 was the pioneer member and had drawn most attention in cancer development in the past decade due to its diverse biological functions. However, controversy regarding SIRTuins and cancer exists and is under debate since they could act as either a tumor suppressor or tumor promoter in different contexts [2]. Recently, SIRT7 has been increasingly identified as crucial regulators for a variety of signaling pathways and has joined the debate [11]. Here we summarized the roles of SIRT7 in tumorigenesis and discussed its therapeutic potential in cancer treatment.
SIRT7 as a Metabolic Modulator
SIRTuins (SIRT1-7) have attracted considerable attention as regulators of metabolism over the past decade. A finely tuned regulatory and evolutionarily conserved switches control and maintain metabolic homeostasis which maintains a delicate balance among energy intake, utilization and storage [12]. A lot of signaling mediators including insulin, insulin growth factor 1 (IGF1) and target of rapamycin (TOR; mTOR in mammals), AMP-activated protein kinase (AMPK) and SIRTuins involved this pathway [12-16].
Previously, SIRTuins (SIRT1-4) have been suggested to modulate glucose homeostasis in various kind of tissues such as muscle, WAT, liver and pancreas. Strikingly, SIRT1 has a controversial role in the control of gluconeogenesis by deacetylating different targets. On one hand, SIRT1 can inhibit hepatic glucose production by downregulating CRTC2 (CREB-regulated transcription coactivator 2) acetylation, leading to its degradation and target genes transcription. On the other hand, SIRT1 can also deacetylate FOXO1 [17, 18] and PGC1α, thus activating the gluconeogenic transcriptional programme. In contrast to SIRT1, other SIRTuins such as SIRT2, SIRT3 and SIRT4 are thought to maintain gluconeogenesis during times of energy limitation [12]. In addition to glucose metabolic regulator, SIRT1 and SIRT6 have been suggested as mediators of fatty acid synthesis and fatty acid oxidation [12]. Interestingly, SIRT7 also joined the team.
Recently, SIRT7 has also been shown to regulate hepatic lipid metabolism by targeting the ubiquitin-proteasome pathway [19]. Yoshizawa and colleagues elegantly demonstrated that SIRT7 knockout mice were resistant to high-fat diet-induced fatty liver, obesity, and glucose intolerance, and that hepatic triglyceride accumulation was also attenuated in liver-specific SIRT7 knockout mice [19]. Hepatic SIRT7 upregulated the protein level of TR4/ TAK1 and activated TR4 target genes to increase fatty acid uptake and triglyceride synthesis/ storage. Biochemically, they revealed that the DDB1-CUL4-associated factor 1 (DCAF1)/damagespecific DNA binding protein 1 (DDB1)/cullin 4B (CUL4B) E3 ubiquitin ligase complex interacted with TR4, leading to its degradation, while binding of SIRT7 to the DCAF1/DDB1/CUL4B complex inhibited the degradation of TR4 [19]. Their data thus suggested a role of SIRT7 in metabolic regulation. However, whether these SIRTuins use same target(s)/mechanism to regulate metabolism is still unknown.
SIRT7 as a Deacetylase: Histone and non-Histone Substrates
In addition to its role as a metabolic regulator, SIRT7 also involved in other signaling pathways by targeting histone or nonhistone proteins as its deacetylation substrates.
It was previously reported that selective hypoacetylation of H3K18Ac has been linked to oncogenic transformation, and is associated with aggressive tumor phenotypes and poor prognosis in patients [20-23]. Through genome-wide screening, Matthew F. Barber et.al. revealed that SIRT7 binds to promoters of a specific set of target genes, where it deacetylates H3K18Ac and promotes transcriptional repression. The spectrum of SIRT7 target genes was defined by its interaction with ELK4, and comprises numerous genes with links to tumor suppression. The deacetylation of H3K18Ac by SIRT7 help human cancer cells maintain anchorage- independent growth and escape from contact inhibition, thus suggesting a oncogenic role of SIRT7 in cancer progression [24].
Not only histone but non-histone proteins can also be deacetylated by SIRT7. Olesya Vakhrusheva et. al. found that SIRT7 interacts with p53 and efficiently deacetylates p53 in vitro, which corresponds to hyperacetylation of p53 in vivo and an increased rate of apoptosis in the myocardium of mutant mice. SIRT7- deficient mice undergo a reduction in lifespan and develop heart hypertrophy and inflammatory cardiomyopathy. SIRT7-deficient primary cardiomyocytes show a 200% increase in basal apoptosis and a significantly diminished resistance to oxidative and genotoxic stress, suggesting a critical role of SIRT7 in the regulation of stress responses and cell death in the heart. This supposed that enhanced p53 activity contributes to the heart phenotype in SIRT7 mutant mice [25].
In addition to histone deacetylation, SIRT7 can also directly affects transcription. It was reported that SIRT7 is an activator of RNA polymerase I transcription by interaction with RNA polymerase I (Pol I) as well as with histones [6]. Overexpression of SIRT7 increases Pol I-mediated transcription, whereas knockdown of SIRT7 reverse the phenomenon. Depletion of SIRT7 inhibits cell proliferation and triggers apoptosis. This suggested that SIRT7 is a positive regulator of Pol I transcription and is required for cell viability in mammals. Interestingly, activation of RNA polymerase I transcription by SIRT7 is associated with its deacetylase activity, since inhibition of the catalytic activity results in decreased association of Pol I with rDNA and a reduction of Pol I transcription [6]. However, the direct deacetylation target in this study was not identified.
Recently, SIRT7 was also reported to target PAF53, a subunit of RNA polymerase I (Pol I), for deacetylation [26]. Acetylation of PAF53 at lysine 373 by CBP and deacetylation by SIRT7 modulate the association of Pol I with DNA, hypoacetylation correlating with increased rDNA occupancy of Pol I and transcription activation. SIRT7 is released from nucleoli in response to different stress conditions, leading to hyperacetylation of PAF53 and decreased Pol I transcription. Nucleolar detention requires binding of SIRT7 to nascent pre-rRNA, linking the spatial distribution of SIRT7 and deacetylation of PAF53 to ongoing transcription. The results identified an RNA-mediated mechanism that adapts nucleolar transcription to stress signaling [26]. Thus, PAF53 was identified as the direct deacetylation target of SIRT7 in pol Imediated transcription.
The Role of SIRT7 in Tumorigenesis
The role of SIRTuins in tumorigenesis is controversial. They can serve as either tumor suppressor or oncogene [2]. Recently, SIRT7 joins the debate [11].
SIRT7 expression has been suggested to be used as a prognostic biomarker in HNSCC. Lai and colleagues [27] used real-time qRT-PCR to investigate the expressions of the seven SIRT genes in human HNSCC tissues to assess the changes in cancerous and noncancerous parts and the correlation with different tumor behaviors. They found that the expression levels of SIRT7 as well as other SIRTuins were significantly downregulated in cancerous tissues compared with noncancerous tissues. The expression levels of SIRT1, SIRT2, SIRT3, SIRT5, and SIRT7 showed downregulation in advanced stages in respect to early stages. They concluded that the downregulation of SIRTuins including SIRT7 expression may contribute to the development of cancer and trigger the neoplastic disease to more advanced stages [27]. However, the direct deacetylation target of SIRT7 in this study was not identified and the function of SIRT7 as a tumor suppressor was not deeply investigated. Thus it’s difficult to draw a conclusion that SIRT7 is a tumor suppressor from this study.
SIRTuin family proteins regulate a variety of cellular signaling events that influence genomic stability, metabolism and ageing, dysregulation of which activity may lead to transformation/tumorigenesis [2]. Recently, SIRT7 has been shown to regulate cancer progression by functioning as an NAD1-dependent H3K18Ac deacetylase, which stabilizes the transformation of cancer cells [24]. Mechanistically, SIRT7 binds to the promoters of a specific set of target genes, where it deacetylates H3K18Ac and promotes transcriptional repression. Barber et al. found that the spectrum of SIRT7 target genes is determined by its interaction with ELK4. Their data conclusively demonstrated that SIRT7 function as an oncogene based on the following reasons. Firstly, selective hypoacetylation of H3K18Ac has been linked to oncogenic transformation, and aggressive tumour phenotypes and poor prognosis [20-23]. Their data suggested that deacetylation of H3K18Ac by SIRT7 is necessary for maintaining anchorage-independent growth and escape from contact inhibition. In addition, SIRT7 is necessary for a global hypoacetylation of H3K18Ac associated with cellular transformation by the viral oncoprotein E1A. Lastly, SIRT7 deficiency significantly reduces the tumor size in xenografts mice. Taken together, their data suggested that SIRT7 acts as an oncogene by deacetylating H3K18Ac [24].
SIRT7 expression and activity has been proposed to be inhibited by hsa-miR-125b. Han and colleagues demonstrated that the expression of hsa-miR-125b and SIRT7 were inversely correlated in bladder cancer. Hsa-miR-125b mimic inhibited, whereas hsamiR- 125b inhibitor promoted the expression of SIRT7. Up-regulation of hsa-miR-125b or down-regulation of SIRT7 inhibited proliferation, motility and increased apoptosis. These data suggest that hsa-miR-125b suppresses bladder cancer development via inhibiting SIRT7 [28].
SIRT7 expression was reported to be dysregulated in human hepatocellular carcinoma (HCC) patients [29]. SIRT7 knockdown causes impaired G1/S phase transition and suppresses growth by upregulating p21WAF1/Cip1 and Beclin-1 and downregulating cyclin D1. Moreover, SIRT7 suppression reduced the tumor growth in a mouse xenograft model. Bioinformatics analysis identified that miR-125a-5p and miR-125b suppressed SIRT7 and cyclin D1 expression and induced p21WAF1/Cip1-dependent G1 cell cycle arrest and treatment of HCC cells with 5-aza-20-deoxycytidine or ectopic expression of wildtype but not mutated p53 restored miR-125a-5p and miR-125b expression and inhibited tumor cell growth, suggesting a role of the promoter methylation and p53 regulation in miRNA activity and SIRT7 expression. Finally, mutations in the DNA binding domain of p53 and promoter methylation of miR-125b analysis suggested that SIRT7 plays a oncogenic role in hepatocarcinogenesis [29].
SIRTuins was also proposed to play a role in colorectal cancer development. The SIRT7 protein level significantly correlated with tumor stage, lymph node metastasis, and poor patient survival. Ectopic SIRT7 expression promoted colorectal cancer cell proliferation, colony formation, and motility while SIRT7 knockdown reversed these phenotypes. In SIRT7-overexpressing cells, the mesenchymal markers vimentin and fibronectin were upregulated, and the epithelial markers E-cadherin and β-catenin were downregulated, suggesting an EMT transition in colorectal cells. Mechanistically, SIRT7 enhanced MAPK pathway activity by p- ERK and p-MEK upregulation. [30].
Concluding Remarks
SIRT7 can function as a transcriptional regulator by targeting Pol I [6], however, factors affecting itself transcription are still unknown. Future investigation need to figure out its upstream regulators which affect its transcription.
Recently researchers spend their effort on inhibitor discoveries and clinical application. However, few of them can be applied to clinical trials due to unknown reasons. Since SIRT7 is not only expressed in normal tissue, but also in tumor tissue, targeting SIRT7 in whole organism will cause problem. Future work may try to target its upstream regulator or downstream effect proteins which are uniquely and abnormally expressed in cancer tissue/patients.
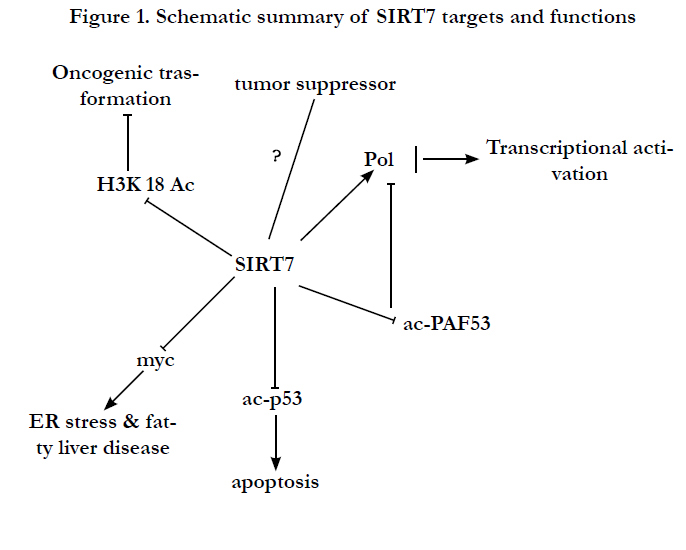
Figure 1. Schematic summary of SIRT7 targets and functions
Acknowledgement
We thank Sinyi Kong for critically reading the manuscript and helpful discussion. This work was supported by the starting fund of ‘One Hundred Plan’ experts from Chongqing University to Zhenghong Lin.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
References
- Kee HJ, Kook H (2011) Roles and targets of class I and IIa histone deacetylases in cardiac hypertrophy. Journal of biomedicine & biotechnology 928326.
- Lin Z, Fang D (2013) The Roles of SIRT1 in Cancer. Genes & cancer 4: 97-104.
- Rauh D, Fischer F, Gertz M, Lakshminarasimhan M, Bergbrede T, et al. (2013) An acetylome peptide microarray reveals specificities and deacetylation substrates for all human sirtuin isoforms. Nature communications 4: 2327.
- Haigis MC, Sinclair DA (2010) Mammalian sirtuins: biological insights and disease relevance. Annual review of pathology 5:253-295.
- Mostoslavsky R, Chua KF, Lombard DB, Pang WW, Fischer MR, et al. (2006) Genomic instability and aging-like phenotype in the absence of mammalian SIRT6. Cell 124: 315-329.
- Ford E, Voit R, Liszt G, Magin C, Grummt I, et al. (2006) Mammalian Sir2 homolog SIRT7 is an activator of RNA polymerase I transcription. Genes & development 20: 1075-1080.
- Tanno M, Sakamoto J, Miura T, Shimamoto K, Horio Y (2007) Nucleocytoplasmic shuttling of the NAD+-dependent histone deacetylase SIRT1. The Journal of biological chemistry 282: 6823-6832.
- Li X, Kazgan N (2011) Mammalian sirtuins and energy metabolism. International journal of biological sciences 7: 575-587.
- Verdin E, Hirschey MD, Finley LW, Haigis MC (2010) Sirtuin regulation of mitochondria: energy production, apoptosis, and signaling. Trends in biochemical sciences 35: 669-675.
- Kiran S, Chatterjee N, Singh S, Kaul SC, Wadhwa R, et al. (2013) Intracellular distribution of human SIRT7 and mapping of the nuclear/nucleolar localization signal. The FEBS journal 280: 3451-3466.
- Li L, Bhatia R (2013) The controversial role of Sirtuins in tumorigenesis - SIRT7 joins the debate. Cell research 23: 10-12.
- Houtkooper RH, Pirinen E, Auwerx J (2012) Sirtuins as regulators of metabolism and healthspan. Nature reviews. Molecular cell biology 13: 225-238.
- Zoncu R, Efeyan A, Sabatini DM (2011) mTOR: from growth signal integration to cancer, diabetes and ageing. Nature reviews. Molecular cell biology 12: 21-35.
- Canto C, Auwerx J (2010) AMP-activated protein kinase and its downstream transcriptional pathways. Cellular and molecular life sciences : CMLS 67: 3407-3423.
- Houtkooper RH, Williams RW, Auwerx J (2010) Metabolic networks of longevity. Cell 142: 9-14
- Fontana L, Partridge L, Longo VD (2010) Extending healthy life span--from yeast to humans. Science 328: 321-326.
- Liu Y, Dentin R, Chen D, Hedrick S, Ravnskjaer K, et al. (2008) A fasting inducible switch modulates gluconeogenesis via activator/coactivator exchange. Nature 456: 269-273.
- Frescas D, Valenti L, Accili D (2005) Nuclear trapping of the forkhead transcription factor FoxO1 via Sirt-dependent deacetylation promotes expression of glucogenetic genes. The Journal of biological chemistry 280: 20589-20595.
- Yoshizawa T, Karim MF, Sato Y, Senokuchi T, Miyata K, et al. (2014) SIRT7 controls hepatic lipid metabolism by regulating the ubiquitin-proteasome pathway. Cell metabolism 19: 712-721.
- Ferrari R, Pellegrini M, Horwitz GA, Xie W, Berk AJ, et al. (2008) Epigenetic reprogramming by adenovirus e1a. Science 321: 1086-1088.
- Horwitz GA, Zhang K, McBrian MA, Grunstein M, Kurdistani SK, et al. (2008) Adenovirus small e1a alters global patterns of histone modification. Science 321: 1084-1085.
- Manuyakorn A, Paulus R, Farrell J, Dawson NA, Tze S, et al. (2010) Cellular histone modification patterns predict prognosis and treatment response in resectable pancreatic adenocarcinoma: results from RTOG 9704. Journal of clinical oncology : official journal of the American Society of Clinical Oncology 28: 1358-1365.
- Seligson DB, Horvath S, McBrian MA, Mah V, Yu H, et al. (2009) Global levels of histone modifications predict prognosis in different cancers. The American journal of pathology 174: 1619-1628.
- Barber MF, Michishita-Kioi E, Xi Y, Tasselli L, Kioi M, et al. (2012) SIRT7 links H3K18 deacetylation to maintenance of oncogenic transformation. Nature 487: 114-118.
- Vakhrusheva O, Smolka C, Gajawada P, Kostin S, Boettger T, et al. (2008) Sirt7 increases stress resistance of cardiomyocytes and prevents apoptosis and inflammatory cardiomyopathy in mice. Circulation research 102: 703-710.
- Chen S, Seiler J, Santiago-Reichelt M, Felbel K, Grummt I, et al. (2013) Repression of RNA polymerase I upon stress is caused by inhibition of RNAdependent deacetylation of PAF53 by SIRT7. Molecular cell 52: 303-313.
- Lai CC, Lin PM, Lin SF, Hsu CH, Lin HC, et al. (2013) Altered expression of SIRT gene family in head and neck squamous cell carcinoma. Tumour biology : the journal of the International Society for Oncodevelopmental Biology and Medicine 34: 1847-1854.
- Han Y, Liu Y, Zhang H, Wang T, Diao R, et al. (2013) Hsa-miR-125b suppresses bladder cancer development by down-regulating oncogene SIRT7 and oncogenic long noncoding RNA MALAT1. FEBS letters 587(23):3875-82.
- Kim JK, Noh JH, Jung KH, Eun JW, Bae HJ, et al. (2013) Sirtuin7 oncogenic potential in human hepatocellular carcinoma and its regulation by the tumor suppressors MiR-125a-5p and MiR-125b. Hepatology 57: 1055- 1067.
- Yu H, Ye W, Wu J, Meng X, Liu RY, et al. (2014) Overexpression of sirt7 exhibits oncogenic property and serves as a prognostic factor in colorectal cancer. Clinical cancer research : an official journal of the American Association for Cancer Research 20: 3434-3445.





