Effect of Substrate Dependent Mushroom Polysaccharide Extracts on IL-7 Up Regulation and Cancer Immunotherapy and, their Probable Mode Of Action
Mwitari, Peter Githaiga1,2, Bian, Yuhong1, Ayeka, Peter Amwoga1,3, Zhang, Yan Jun1.
1. Tianjin University of Traditional Chinese Medicine, 312 Anshan West Road, Nankai District, Tianjin, P.R. China, 300193.
2. Kenya Medical Research Institute. P.O. Box 54840-00200, Nairobi, Kenya.
3. Egerton University, Kenya
*Corresponding Author
Mwitari, Peter Githaiga,
Kenya Medical Research Institute,
P.O. Box 54840-00200, Nairobi, Kenya.
E-mail: pmwitari67@yahoo.com
Article Type:Research Article
Received: August 08, 2014; Accepted: August 27, 2014; Published: August 28, 2014
Citation: Mwitari PG, Bian Y, Ayeka PA, Zhang YJ (2014) Effect of substrate dependent mushroom polysaccharide extracts on IL-7 up regulation and cancer immunotherapy and, their probable mode of action. Int J Cancer Stud Res. 3(3), 41-57. doi: dx.doi.org/10.19070/2167-9118-140007
Copyright: © 2014 Monica C S. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
Ethnopharmacological relevance:
Fungi have been used in traditional systems both as food and medicine. In China the mushrooms are widely used in Traditional Chinese Medicine (TCM). On one hand residue resulting from medicinal plants are treated as waste, yet the environment in which medicinal plants including fungi grow greatly influences the potency and quantities of bioactive secondary metabolites thereof. Since Mushrooms are widely consumed for immunomodulatory purposes especially for people suffering from tumors, this study sought to find out whether there is enhanced effect of mushroom polysaccharides extracts of Trametes robiniophila murr a Chinese fungus, grown on media supplemented with Chinese herbal drug residues both in vitro and in vivo as an immunotherapeutic remedy for cancer.
Methodology:
We used Intestinal Epithelial Cells (IEC) known to produce IL-7 cytokine which works on the immune system and on the postulation that polysaccharide extracts work by up regulating IL-7. The polysaccharide extracts investigated were obtained from mushrooms grown on media supplemented with Chinese herbal residues. The Cell Counting Kit-8 (CCK-8) proliferation assay procedure was used to asses’ cell proliferation. Extract concentrations ranging from 0.00001 - 100µg/ml was used in the cell proliferation assay. IEC-6 cells grown in media supplemented with 100µg/ml mushroom polysaccharides extracts was used in vitro and RT-PCR technique employed to evaluate up-regulation of IL-7 in a time dependent manner ranging from 3 to 72 hours. The extract with best IL-7 expression was selected for further evaluation in vivo at 30mg/mice/day in mice infected with CT-26 colon cancer tumor cells. Cyclophosphamide at 20mg/mice/day was taken as the positive control drug. Flow cytometry was employed to evaluate the peripheral blood CD4+ and CD8+ lymphocytes. The extract was further tested in vitro on CT-26 cells (100 & 200 µg/ml), to explore direct inhibition.
Results:
1µg/ml was found to give optimum IEC-6 cell growth however; 100µg/ml was determined as the ED50 and used as the minimum evaluation concentration for IL-7 expression. The polysaccharide extracts obtained from mushrooms supplemented with the Chinese herb Gancao (Glycyrrhiza glabra L. var. glandulifera) residues were found to up-regulate IL-7 best. 6 hours exposure of cells to extract gave optimal IL7 expression of over 1.2 times at 100µg/ml compared to polysaccharide extracts from mushrooms grown on normal media. Extracts obtained at temperatures ranging between 70-90°C were found to be most ideal. The extract was found to increase the CD4+ and CD8+ lymphocytes in vivo in mice a reflection on IL-7 up regulation in the intestinal mucosa and further to inhibit CT-26 cell growth in a significant way at P≤0.05.
Conclusion:
Polysaccharide extracts obtained from mushroom Trametes robiniophila murr grown on media supplemented with Chinese herb Gancao (G. glabra L. var. glandulifera) residues offers increased immunomodulatory effect as an alternative treatment to cancer cases. It can be employed as a prescription supplement for immunotherapy and immuno surveillance. The mechanism of action is probably stimulation of IEC-6 cells to produce increased IL-7 Cytokine responsible for resultant adaptive immunity and cancer immuno surveillance. The polysaccharide extract too has chemotherapeutic effect and is safe.
2.Background
3.Methodology
3.1.Cells and extract
3.2.RT-PCR assay technique
3.3.In vivo studies of selected Mushroom Polysaccharide extracts
3.4.Data analysis
4.Results
4.1.Cell Proliferation assay
4.2.Expression of GAPDH and IL-7 by Mouse IEC-6 cells subjected to various extracts at a concentration of 100μg/ml
4.3.Mouse IEC-6 cells subjected to 20 Mushroom Polysaccharide extracts extracted at varying Temperatures and tested for their effect on the expression of GAPDH and IL-7 mRNA at 3.33mg/ml exposed for 72 hours
4.4.Time and Dose Dependent In Vitro Studies of IEC-6 Cells Expression of IL-7 mRNA on Subjection to Media Supplemented with Polysaccharide Extracts from Substrate Dependent Mushrooms
4.5.Mean Weight Indexes of Mice Tumor and Immune Organs
4.6.Flow Cytometry- Immunofluorescence staining of mice Cells by Lysed Whole Blood Method
4.7.Relative IL 7 Gene expression in mice small intestines
4.8. Mouse Interleukin 7 cytokine (IL 7) in serum and Intestinal mucosa
4.9.Levels of other Cytokines in Peripheral blood
5.Discussion
6.Conclusion
7.Acknowledgement
8.References
Key Words
Mushroom polysaccharides extracts, Trametes robiniophila murr, IL-7 Cytokine, CT-26 colon cancer tumor cells, CD4+ and CD8+ lymphocytes, immunity.
Background
The world has witnessed an upsurge in cancer cases, a disease that afflicts both men and women worldwide. Death due to cancer especially is projected to rise to over 11million in 2030 [1]. In China, cancer has become the leading cause of death. Mortality related to cancer in China has risen by 80% during the last 30 years to an alarming number of 1.8 million cancer deaths in 2010 [2]. During the last three decades, deaths related to lung cancer have increased by 465% [2]. China alone contributes more than half of the world’s newly diagnosed liver and esophageal cancer cases, and 42% of newly diagnosed stomach cancer cases [3]. The disease burden due to cancer cannot be overemphasized.
Cancer is by and large associated with our body’s immune system.Concerted efforts in addressing the health challenge due to cancer, by, identifying possible solutions towards reduction and treatment are greatly desired. Some medicinal plants and their components are known to boost immunity. Biological properties of medicinal mushrooms have been investigated for decades; especially for antitumor and immunostimulatory effects of their polysaccharide compounds [4]. Mushrooms exhibit anti-tumor activity based on the modulation of immune system and this activity is demonstrated for crude fungal extracts and isolated compounds like polysaccharides, polysaccharo-peptides, polysaccharide-proteins and proteins from culture media of various mushrooms[5]. Their polysaccharide fraction which is mainly composed of β- glucans responsible for immune modulating effects by a number of ways such as activating leukocytes which are depended on the structural characteristics of the β- glucans, more so, they contain different biological active compounds like small proteins, lectins, and polyphenols which are mainly β- glucans present in the cell walls [5]. It has been reported that this β-glucans increase the phagocytic activity and production of reactive oxygen intermediates, inflammatory mediators and cytokines by leukocytes [6,7]. More than 50 mushroom species have yielded potential immunoceuticals with immunomodulatory and antitumor effects, they include lectines, polyssacharides, polysaccharides-peptides, polyssacharide-proteins complexes like lentinan, schizophyllan, polysaccharide-K, polysaccharide p, active hexose correlated compounds, AHCC, maitake D fraction which are regarded as biological response modifiers (BRM) and have been investigated in human cancer [7,8]. Pleurotus ostreatus has anti-cancer, immunomodulatory, antiviral, antibiotic, and anti-inflammatory and cholesterol lowering activities [8]. The water extract from its mycelia and fruit bodies have a role in increasing production of ROS from neutrophils and also has immunomodulatory properties on all immunocompetent cells [8].
Ganoderma lucidium is known for longetivity-promoting and as tonic herb and, the biological activities of Ganoderma lucidium especially its anti-tumor and immunomodulatory properties show that its extract is able to stimulate the inflammatory response due to the expression of IL-1, IL-6, and TNF-α linked to the antitumor activity[9,10]. Isolated polysaccharides and triterpenoids from this mushroom are potent inhibitors of in vitro and in vivo tumor growth [11]. Trametes robiniophila murr has been applied in TCM for approximately 1600 years [12] though recently its anticancer properties have been put into complementary therapy and its water extracts are proteoglycans. This extract is able to display apoptosis, antiangiogenesis, drug resistance reversal, and antimetastasis and system immune activation [13]. Botanical polysaccharides thus provide a unique opportunity for the discovery of novel therapeutic agents that exhibit beneficial immunomodulatory properties.
IL-7 has a potential for adoptive immunotherapy [14]. Cellular immunity against tumors initially focused on CTLs, but in recent years it has become evident that CD4+ T cells also play a critical role in the development of effective anti-tumor immunity [15]. IL-7 is implicated in synthesis and survival of CD4+ lymphocytes [15] Furthermore, IL-7 has the potential to induce cytotoxic T cells in lung metastases of murine sarcomas [14]. In addition, it has been shown to induce lymphokine-activated killer cell activity obtained from patients early after bone marrow transplantation [14]. Adoptive transfer of tumor-reactive T cells into lympho-depleted hosts has shown that a significant percentage of treated patients can experience immune-mediated cancer regression across multiple institutions [16,17,18].
IL-7 prolongs survival, decreases tumor burden and enhances tumor T cell infiltration in vaccinated mice [19,20]. Histology of tumors from IL-7–treated mice is heavily infiltrated with both CD4+ and CD8+ T cells[19,20]. The presence of tumor infiltrating lymphocytes (TILs) within the tumor microenvironment is considered to be an indication of the host immune response to tumor antigens [19], and is thought to reflect the dynamic process of “cancer immunoediting”.
Intestinal epithelial cells (IEC) have been implicated in IL-7 synthesis [21]. IL-7 plays an important role in immune processes in our bodies. Several studies have indicated that IEC may play an important role in mucosal immune responses by helping to regulateintestinal intraepithelial lymphocytes (IEL) [22]. Interleukin-7 has been implicated and proposed as an important link to finding the right treatment for cancer ailments. IL-7 has a potential for adoptive immunotherapy [23,24,25,26,27,28]. Importance and usefulness of Cytokine IL 7 as a tool in cancer treatment research has been demonstrated before [29]. In the current study, it is hypothesized that Polysaccharide extracts obtained from mushrooms grown in media supplemented with Chinese herbal residues, act on the intestinal mucosal cells (IEC) which results in up regulation of the production of IL-7. IL-7 then elicits cascade events boosting immune response to cancer cells.
Drug agents that enhance or stimulate the production of IL-7 therefore provide potential candidates in cancer treatment. IL-7 too is a viable research tool in evaluation of potential plant medicines and their mode of action
Methodology
Mouse IEC-6 cells were used to investigate effects of mushroom polysaccharide extract obtained from mushrooms grown on media supplemented with known quantities of Chinese medicinal plant residues of Gancao (Radix Glycyrrhizae grabra L.) and Huangqi (Astragalus membranaceus (Fisch) Bunge). The standard cell culture techniques were employed and media added with extracts of known concentrations at specified time frames. The Chinese mushroom investigated was Huaier (Trametes robiniophila murr). The Mushroom species was botanically identified by a plant taxonomist at TUTCM and grown at alternating 12 hour cycles of light and dark in culture bottles. Water soluble mushroom polysaccharides were extracted as described by Mizuno [30]. The extraction conditions were optimized at 70-90°C in water, extract freeze dried and stored at 4°C until use.
IEC-6 cells (ATCC) were seeded at 50,000 cells per well in corning 96 well flat bottomed micro titer plates and incubated overnight at a volume of 100µl. 10µl mushroom substrates (0680, 0790 & M) at eight different concentrations each serially diluted were added and cells incubated for a further 72hours. 10µl of CCK-8 (Dojindo Molecular Technologies, Beijing,China) was then added and incubated in a high humidity environment at 37°C and 5% CO2 for 3 hours and optical density (OD) read at 460 nm in a 96-well micro titer plate Tecan i-control infinite 200 OD reader. The test was done in triplicate. The data was analyzed using unpaired Student's T-test. Values are expressed as mean ± S.D.
A second assay was carried out to determine the reliability and cell numbers by plating 100µl of a known number of cells in a clear 96 well plate (Cell numbers ranging from 2x104-2x106). 10 µl CCK-8 solution was then added and incubated for 3 hours under identical conditions to first assay and absorbance determined.
CT-26 colon cancer cells (ATCC) were seeded at 50,000 cells per well in corning 96 well flat bottomed micro titer plates and incubated overnight at a volume of 100µl. Mushroom polysaccharide extract was tested at two concentrations of 100 & 200µg/ml by adding 10µl of each and cells incubated at 37°C, 5% CO2 high humidity for a further 72hours. Two controls were similarly set up; a negative control without drug and a positive control where Cytoxin at 100µg/ml was added. 10µl of CCK-8 (Dojindo Molecular Technologies, Beijing, China) was then added and incubated in a high humidity environment at 37°C and 5% CO2 for 3 hours and optical difference (OD) read at 460 nm in a 96-well microtiter plate Tecan i-control infinite 200 OD reader. The test was done in triplicate. The data was analyzed using unpaired Student's T-test. Results are presented as mean ± S.D and in form of a bar chart.
In this study polysaccharide bearing extracts from mushrooms grown on normal media supplemented with Chinese herbal residues were investigated on IEC-6 cells in vitro. The levels of expression of IL-7 mRNA were determined by RT-PCR methods as a measure of their immune modulating enhancement effect. Appropriate controls were set up with polysaccharide extracts from mushrooms grown on normal media only.
Extraction and amplification was done according RNeasy® Mini Kit (Qiagen).
The IEC-6 cells were cultured in 6 well plates in media supplemented with various extracts at concentrations of 100µg/ml for 72, 6 or 3 hours. Briefly the media was decanted and cells washed in PBS (D-hanks solution) and 1ml Trizol (Invitrogen) added. 0.2ml of Chloroform at 4°C was added per 1ml shaken vigorously using a vortex for 30 seconds and incubated at room temperature (about 20°C) for a period of 4 minutes. This was followed by centrifuging for 15 minutes at 12000 revolutions per minute (rpm). A colorless aqueous layer formed at the top of which 500µl was pipetted out carefully. A similar volume (500µl) of isopropyl alcohol at minus 20°C was added to the RNA fraction and vortexed properly and incubated at room temperature (about 20°C) for a period of 25 minutes. The mixture was centrifuged for 10 minutes at 12000 revolutions per minute (rpm) at 4°C. The supernatant was discarded and RNA pellet washed with 1ml of 75% ethanol and vortexed properly and, centrifuged for 10 minutes at 8000 revolutions per minute (rpm) at 4°c. A refrigerated centrifuge was used each time. The supernatant was discarded and RNA pellet air dried before re-dissolving in 20 µl RNAse free water. The Optical difference (OD) was measured at 260/280nm to determine the quality of RNA using a NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies, Inc., Wilmington, DE, USA) and concentration (ng/µl) obtained. The concentration of RNA (ng/µl) was used to calculate the volumes of RNA and water (H2O) for use in reverse transcription by first dividing 500 by ng/µl of RNA obtained to give volume of RNA in µl and then this was subtracted from 6.5 to give volume of H2O to be used. Reverse transcription and cDNA amplification was done according to RNeasy ® Mini Kit (Qiagen), and the following genes were targeted for amplification; GAPDH with primers sequence 5’ to 3’ sense ACC ACA gTC CAT gCC ATC AC and antisense TCC ACC ACC CTg TTg. CTg TA and, IL-7 sense gAg TTT CAg ACg gCA CAC AA and antisense gAA ACT TCT ggg Agg gTT CC (from Sangon co.ltd) at reaction conditions (94°C for 3 minutes, 94°C for 30 seconds, 60.5°C for 30 seconds, 72°C for 30 seconds, 4°C indefinitely) and at 22 cycles for GAPDH and 38 cycles for IL-7. All amplified cDNA were analyzed by 1.5% agarose gel electrophoresis and stained with ethidium-bromide for visualization under UV illumination (GeneGenius) and photographed. The size of amplified DNA was identified by comparison with DNA marker (100 bp DNA ladder, TaKaRa Biotechnology (Dalian) co., Ltd) and volumes of the DNA calculated for each band.
45 female Balb/C mice 6 weeks old weighing on average 18.2g and infected with CT-26 Colon cancer tumor cells were employed to assess the immune boosting action of mushroom polysaccharide extracts. They were housed in an animal facility well lighted by natural light and well ventilated. They were kept in groups of five per cage. They received normal mice feed and water ad libitum. The experimental period was 17 days with two days to acclimatize and one day after 14 days of treatment. The Animal experiment was carried out in accordance with Tianjin University of traditional Chinese medicine guidelines on laboratory animal’s use and care.
The mice were divided into 3 groups of 15 mice each in a completely randomized design consisting of a negative control which received 0.2ml normal saline orally, a positive control group which received 20mg/ml of an anti-cancer drug Cyclophosphamide (Cytoxin) through IP route and, a test group which received 30mg of mushroom polysaccharides extract orally.
The treatments were administered on a daily basis for two weeks except Cytoxin which was administered for 10 days. Weights of the animals were taken at 0 days, 10 days, and 15th day before they were sacrificed and blood, tumor, liver, spleen, thymus and lungs excised and properly labeled. The blood was collected in 0.4ml of 6mg/ml EDTA to prevent coagulation and stored at 4 degrees centigrade. The organs were weighed and either stored at minus 20 or minus 80 degrees centigrade and histopathology evaluation done. Lysed Whole blood immunofluorescence staining was performed.
Whole blood (peripheral) was drawn from the eyes into 1.5 ml centrifuge tubes. The tubes were pre-coated with 0.4ml of 6mg/ ml EDTA as an anticoagulant. The tubes were well rinsed with the 0.4ml of EDTA by corking and swilling round to cover all sides of the tube before opening to dry the sides and collecting the blood. All tubes were properly labeled. Briefly, 100µl of wellmixed anti-coagulated whole blood was added to the bottom of a well labeled tube. The appropriate primary antibody was then added to each tube as described in the following manner, to 100ul (1x106) Cells was added 0.25µg of FITC Hamster Anti-Mouse CD3e, 0.25µg of APC Rat Anti-Mouse CD4 and 1.0µg of PE Rat Anti-MouseCD8a. The tube was capped and vortexed gently to mix and, incubated for 25 minutes in the dark at room temperature (20° to 25°C). The tube was removed from dark chamber and mixed well. 1.5ml of lysing solution was added to and vortexed. After which it was incubated at room temperature in the dark for 10-15 minutes, until cells become clear and centrifuged for 5 minutes at 1000 rpm (200 x g). The supernatant was removed by aspiration, vortexed and 2 ml washing solution added and, centrifuged for 5 minutes at 1000 rpm (200 x g). The supernatant was again removed by aspiration. The cells were fixed by re-suspending in 2% paraformaldehyde buffer for 30 minutes at room temperature, and washed with wash buffer. The cells were re-suspended in 500 µl wash buffer and stored in the refrigerator at 4°C for 24 hours in the dark before reading in a Merck- Millipore flow cytometer. This exercise was repeated for all samples. A negative control was set up in one tube by titrating 50ul of cells and adding 100ul PBS and taken through the same procedure.
Statistical analysis was done using excel d ata sheets and SPSS version 14.0. The expression of IL-7 mRNA relative to GAPDH mRNA was calculated and bar charts drawn. The differences between the control and the treatments in these experiments were tested for statistical significance by unpaired Student's T-test. A value of P<0.05 was considered to indicate statistical significance. Values are expressed as mean ±S.D.
Results
A calibration chart was drawn to ascertain reliability and help enable cell number determination from absorbance values. The cell numbers were observed to have a linear relationship with the absorbance values. A high number of cells gave a corresponding high absorbance. A concentration of 1µg/ml gave the best proliferative effect on IEC-6 cells for the all tested extracts (Table1, Figure 1). Mushroom extracts investigated had no visible cytotoxic effects and promoted cell growth normally even at concentrations above 100µg/ml in 6 well plates. The ED50 was determined to be approximately 100 µg/ml.
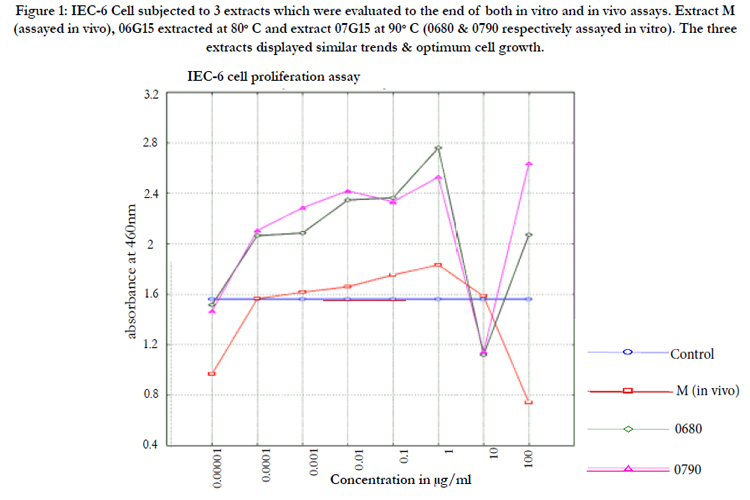
Figure 1: IEC-6 Cell subjected to 3 extracts which were evaluated to the end of both in vitro and in vivo assays. Extract M (assayed in vivo), 06G15 extracted at 80°C and extract 07G15 at 90°C (0680 & 0790 respectively assayed in vitro). The three extracts displayed similar trends & optimum cell growth.
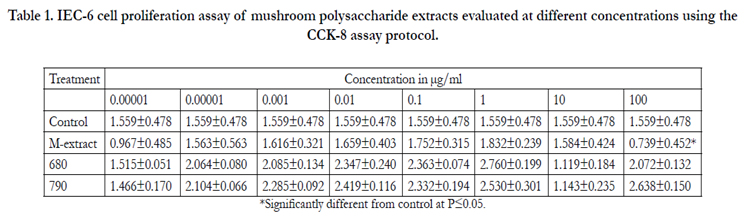

Table 1. IEC-6 cell proliferation assay of mushroom polysaccharide extracts evaluated at different concentrations using the CCK-8 assay protocol.
*Significantly different from control at P≤0.05.
IEC-6 cells were shown to express IL-7 Cytokine constitutively.This cytokine is associated with immunomodulation and cancer cells Immunosurveillance. IEC- 6 cells subjected to mushroom polysaccharide extracts coded as; 01C, 01G15, 01H15, 03C,
03G15, 03H15, 06C, 06G15, 06H15, 07C, 07G15 & 07H15. These were water extracts from Huaier (Trametes robiniophila murr) mushrooms grown in media supplemented with chinese herbs Gancao (G) and Huangqi (H) and extracted at varying conditions to determine the right herbal supplement and optimum extraction conditions. They were tested in IEC-6 cells for IL-7 mRNA expression as a measure of their bioactivity ( Figures 2A & B). Extracts 06G15 and 07G15 were selected as having the best IL-7 mRNA expression effects.
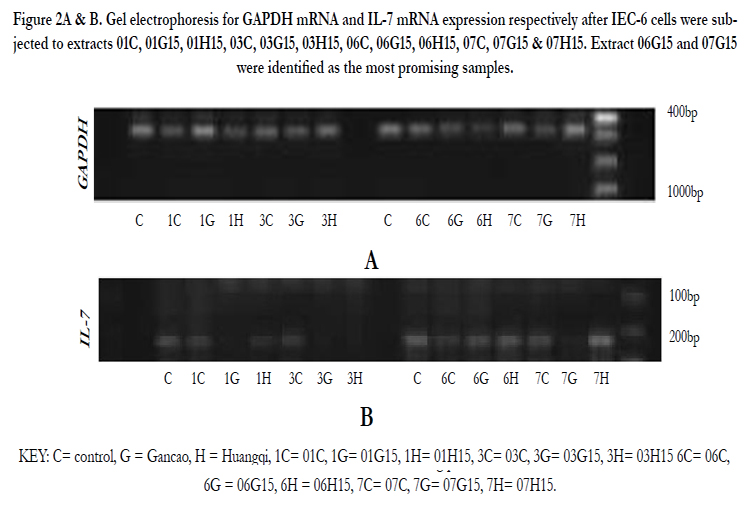
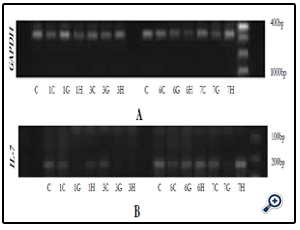
Figure 2A & B. Gel electrophoresis for GAPDH mRNA and IL-7 mRNA expression respectively after IEC-6 cells were subjected to extracts 01C, 01G15, 01H15, 03C, 03G15, 03H15, 06C, 06G15, 06H15, 07C, 07G15 & 07H15. Extract 06G15 and 07G15 were identified as the most promising samples.
KEY: C= control, G = Gancao, H = Huangqi, 1C= 01C, 1G= 01G15, 1H= 01H15, 3C= 03C, 3G= 03G15, 3H= 03H15 6C= 06C, 6G = 06G15, 6H = 06H15, 7C= 07C, 7G= 07G15, 7H= 07H15.
Extracts 06G15 and 07G15 were selected as best in up regulating IL-7. The extraction of the mushroom polysaccharides was again done at varying temperatures and IL-7 mRNA expression used to select the optimum extraction temperatures. 06G15 at 60, 70 & 80°C and, 07G15 at 90°C were found to have the best effects on IL-7 mRNA expression of 1.4 or more times (Figures 3A & B) compared to the control and the rest of the extracts.
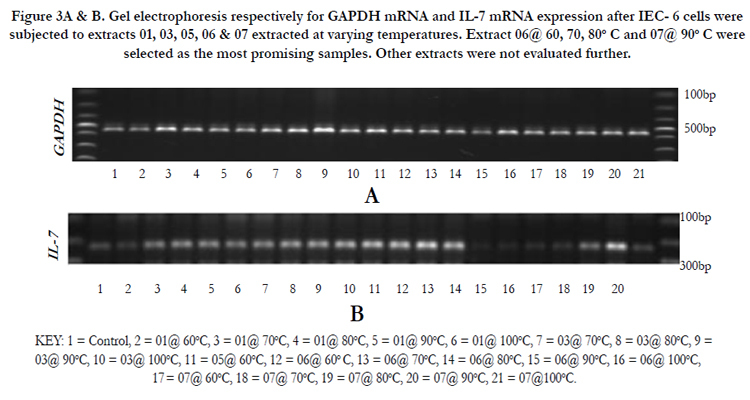
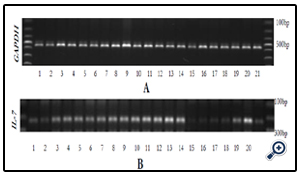
Figure 3A & B. Gel electrophoresis respectively for GAPDH mRNA and IL-7 mRNA expression after IEC- 6 cells were subjected to extracts 01, 03, 05, 06 & 07 extracted at varying temperatures. Extract 06@ 60, 70, 80°C and 07@ 90°C were selected as the most promising samples. Other extracts were not evaluated further.
KEY: 1 = Control, 2 = 01@ 60°C, 3 = 01@ 70°C, 4 = 01@ 80°C, 5 = 01@ 90°C, 6 = 01@ 100°C, 7 = 03@ 70°C, 8 = 03@ 80°C, 9 =03@ 90°C, 10 = 03@ 100°C, 11 = 05@ 60°C, 12 = 06@ 60° C, 13 = 06@ 70°C, 14 = 06@ 80°C, 15 = 06@ 90°C, 16 = 06@ 100°C, 17 = 07@ 60°C, 18 = 07@ 70°C, 19 = 07@ 80°C, 20 = 07@ 90°C, 21 = 07@100°C.
IEC-6 cells on exposure to mushroom (Huaier) extracts at two concentrations of 100µg/ml and 3.33mg/ml were found to promote the expression of IL-7 mRNA positively (Figures 4A & B, 5A & B). The cells were exposed to 6 hours and 3 hours to gauge IL-7 mRNA expression at the two concentrations in a time dependent way. The expression at 3 hours was not significantly different from the control (results not provided) but a significant difference is noticed on exposure to 6 hours. At 100µg/ml the expression of IL-7 mRNA is better (Table 3, Figure 5C) compared to the higher concentration of 3.33mg/ml (Table 2, Figure 4C). IL-7 mRNA expression is significantly different for extract 0680 at 3.33mg/ml and 0670 at 100 µg/ml from the extract from mushrooms grown on normal media (Tables 2&3). The extracts under investigation express IL-7 mRNA approximately 1.2 - 2 times more compared to the house keeping gene GAPDH mRNA (Tables 2&3).
Figure 4A & B, Table 2, Figure 4C

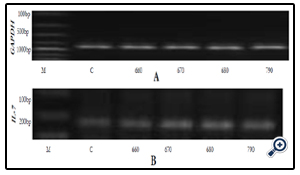
Figure 4A & B. Gel electrophoresis respectively for GAPDH mRNA and IL-7 mRNA expression after IEC- 6 cells were subjected for 6 hours to extracts C, 06G15@60°C, 06G15@70°C, 06G15@80°C & 07G15@90°C at a concentration of 3.33mg/ ml. Extracts obtained at the 4 different temperatures 60, 70, 80 & 90° C respectively.
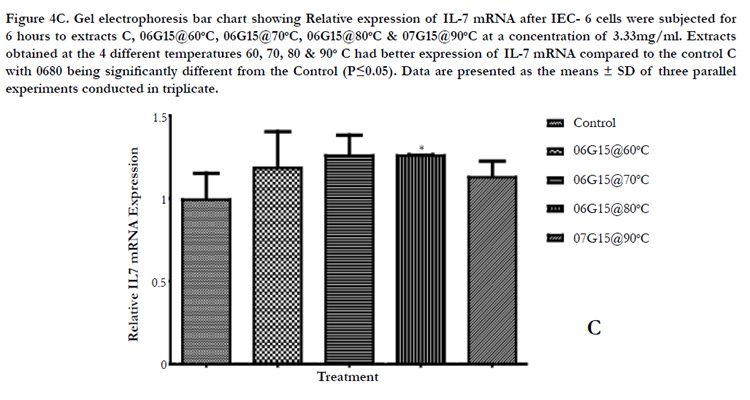
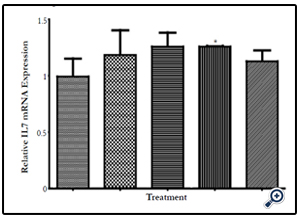
Figure 4C. Gel electrophoresis bar chart showing Relative expression of IL-7 mRNA after IEC- 6 cells were subjected for 6 hours to extracts C, 06G15@60°C, 06G15@70°C, 06G15@80°C & 07G15@90°C at a concentration of 3.33mg/ml. Extracts obtained at the 4 different temperatures 60, 70, 80 & 90° C had better expression of IL-7 mRNA compared to the control C with 0680 being significantly different from the Control (P≤0.05). Data are presented as the means ± SD of three parallel experiments conducted in triplicate.
Key: M-Marker, C-Control, Extracts, 660 = O6G15@ 60°C, 670 = O6G15@ 70°C, 680 = O6G15@ 80°C, 790 = O7G15@ 90°C
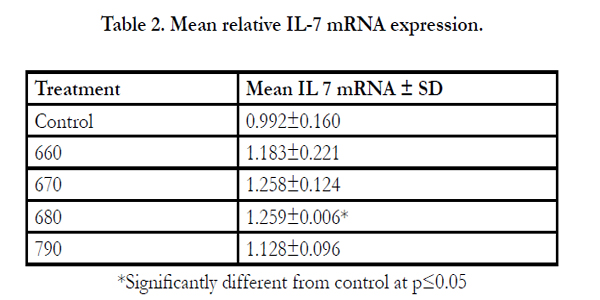
Table 2. Mean relative IL-7 mRNA expression
*Significantly different from control at p≤0.05
Figure 5A & B, Table 3, Figure 5C
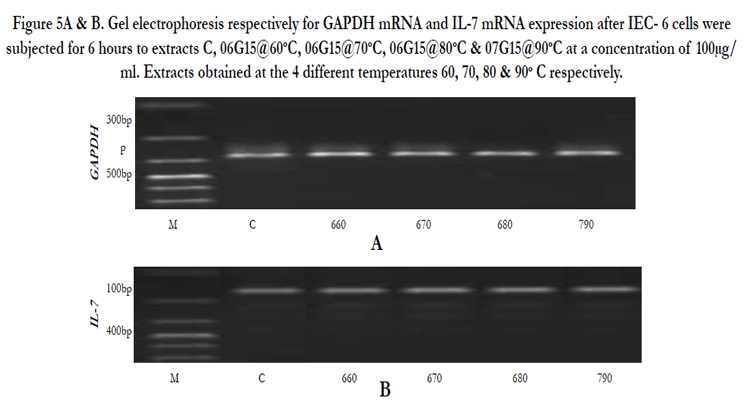
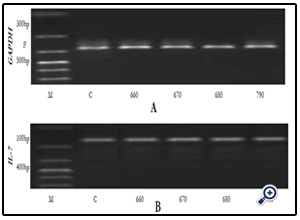
Figure 5A & B. Gel electrophoresis respectively for GAPDH mRNA and IL-7 mRNA expression after IEC- 6 cells were subjected for 6 hours to extracts C, 06G15@60oC, 06G15@70°C, 06G15@80°C & 07G15@90°C at a concentration of 100μg/ml. Extracts obtained at the 4 different temperatures 60, 70, 80 & 90° C respectively.
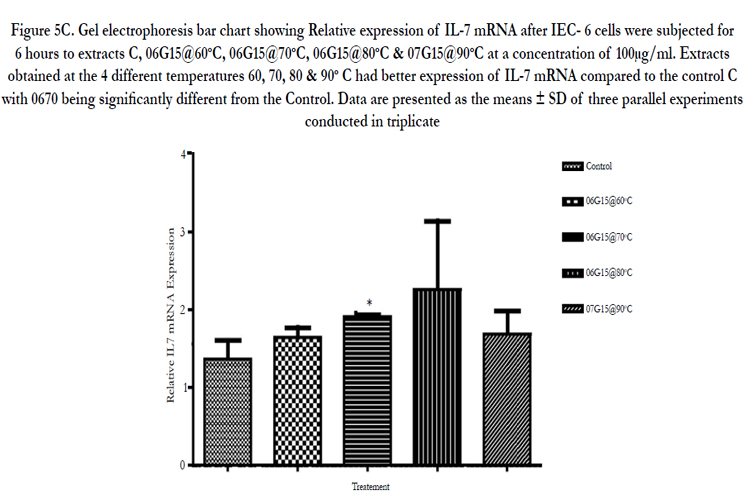
Figure 5C. Gel electrophoresis bar chart showing Relative expression of IL-7 mRNA after IEC- 6 cells were subjected for 6 hours to extracts C, 06G15@60°C, 06G15@70°C, 06G15@80°C & 07G15@90°C at a concentration of 100μg/ml. Extracts obtained at the 4 different temperatures 60, 70, 80 & 90° C had better expression of IL-7 mRNA compared to the control C with 0670 being significantly different from the Control. Data are presented as the means ± SD of three parallel experiments conducted in triplicate.
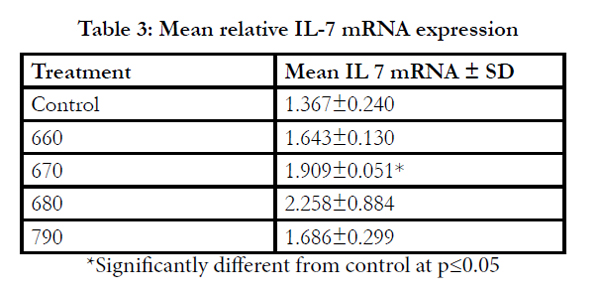
Table 3: Mean relative IL-7 mRNA expression
*Significantly different from control at p≤0.05
After commencing treatment (Figure 6) there was a slight decrease in weight for all groups but the tread reversed and started to increase gradually over the treatment period. The mean tumor weight indices in the polysaccharide fed test group infected with Ct-26 colon cancer cells did show a significant difference at p≤0.05 (Table 4, Figure 7). Conversely the mortality in the negative control (no treatment) group was higher and only about half of the mice 53.3% survived while in the positive control and test groups, the survival rate was over 73% (Table 5, figure 8). The mean weights indices of immunity organs, the Thymus and the Spleen were significantly different from those of the normal uninfected mice. The Thymus weight indexes were significant in a negative way by having lower values and the Spleen weight indexes were significant (p≤0.05) in a positive way by having higher values (Table 6, figure 9 & 10). A larger spleen size may suggest a heightened baseline activation state or pre-expansion of a responding cell population
Figure 6. Balb/C mice infected with CT -26 tumor cells. Figure shows the tumor resulting from the infection.
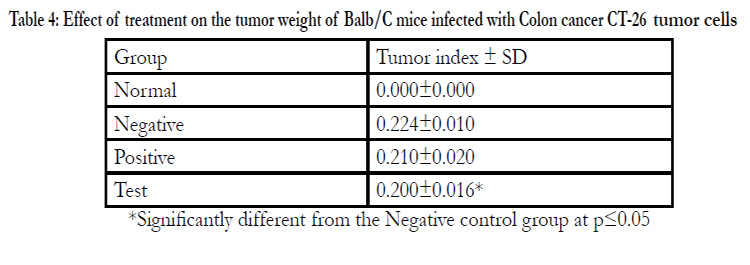
Table 4: Effect of treatment on the tumor weight of Balb/C mice infected with Colon cancer CT-26 tumor cells
*Significantly different from the Negative control group at p≤0.05

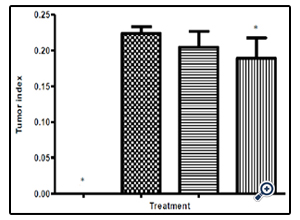
Figure 7: The mean tumor index in the normal group and three experimental groups of Balb/C mice infected with Colon Cancer tumor cells. Test group has a significant difference compared to the untreated negative control group.
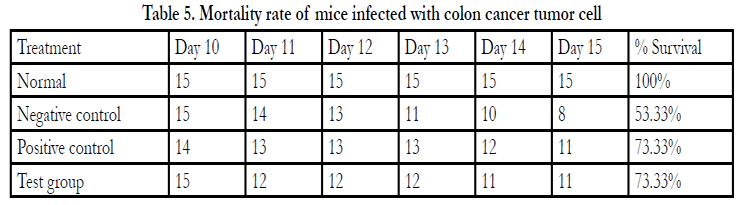
Table 5. Mortality rate of mice infected with colon cancer tumor cell
*Significantly different from the Negative control group at p≤0.05
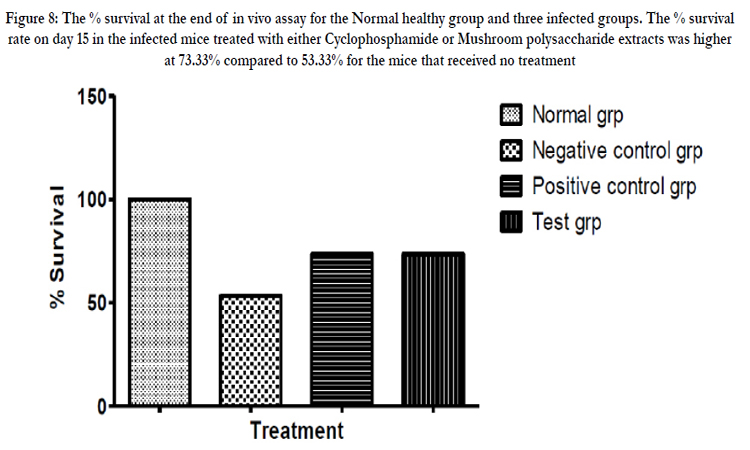
Figure 8: The % survival at the end of in vivo assay for the Normal healthy group and three infected groups. The % survival rate on day 15 in the infected mice treated with either Cyclophosphamide or Mushroom polysaccharide extracts was higher at 73.33% compared to 53.33% for the mice that received no treatment.
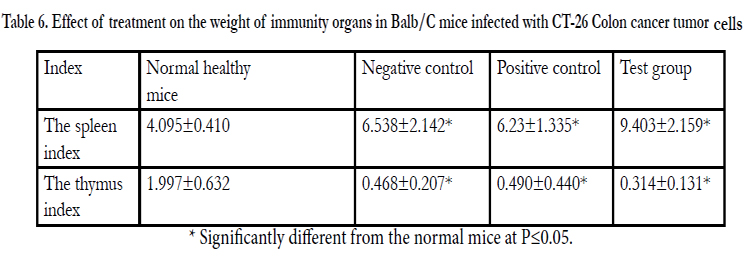
Table 6. Effect of treatment on the weight of immunity organs in Balb/C mice infected with CT-26 Colon cancer tumor cells.
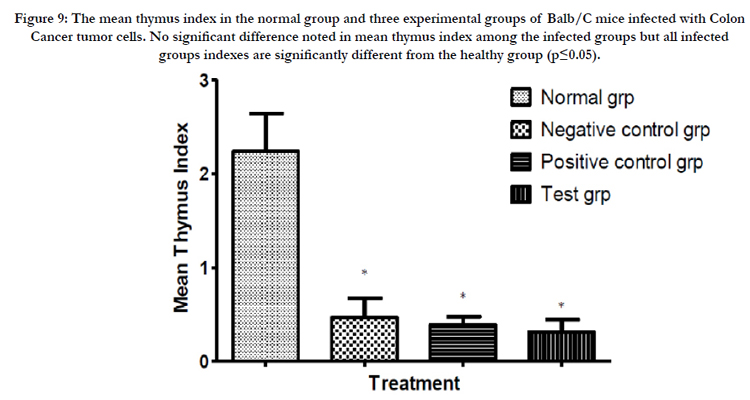
Figure 9: The mean thymus index in the normal group and three experimental groups of Balb/C mice infected with Colon Cancer tumor cells. No significant difference noted in mean thymus index among the infected groups but all infected groups indexes are significantly different from the healthy group (p≤0.05).
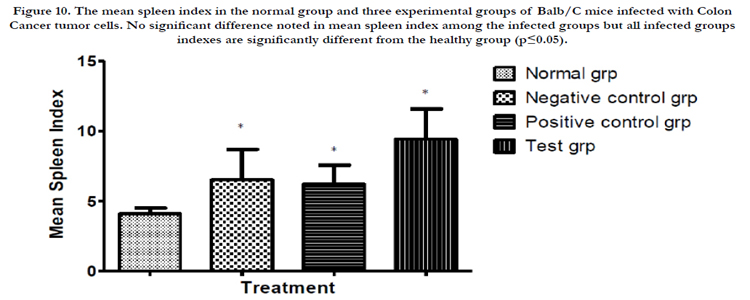
Figure 10. The mean spleen index in the normal group and three experimental groups of Balb/C mice infected with Colon Cancer tumor cells. No significant difference noted in mean spleen index among the infected groups but all infected groups indexes are significantly different from the healthy group (p≤0.05).
Flow Cytometry analysis of whole mouse peripheral blood was done to determine the effect of different treatments on the lymphocytes associated with immunomodulation and cancer cell Immunosurveillance. Total CD3+, CD4+ and CD8+ cell numbers were assessed. The CD4+ and CD8+ cells are represented as a % of the total CD3+ cells. The CD4+ and CD8+ cells in the mice which received the mushroom polysaccharide extracts were found to be significantly different (p≤0.05) from the untreated control mice (Table 7, Figures 11, 12, 13).
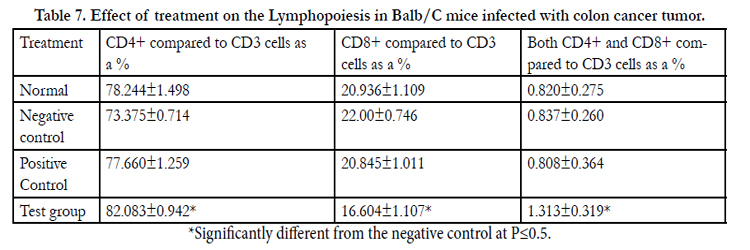
Table 7. Effect of treatment on the Lymphopoiesis in Balb/C mice infected with colon cancer tumor.
*Significantly different from the negative control at P<0.5
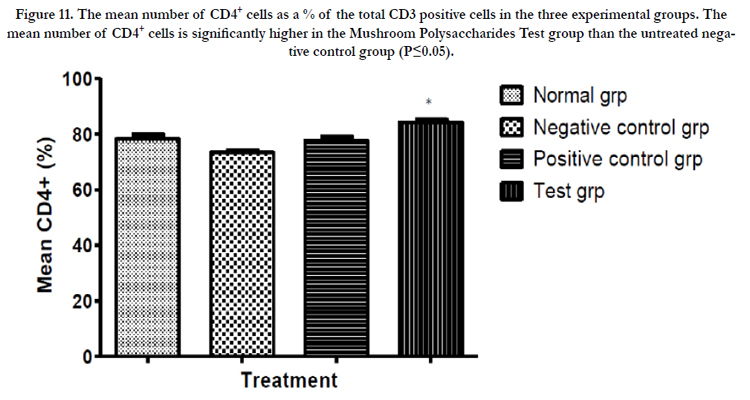
Figure 11. The mean number of CD4+ cells as a % of the total CD3 positive cells in the three experimental groups. The mean number of CD4+ cells is significantly higher in the Mushroom Polysaccharides Test group than the untreated negative control group (P≤0.05).
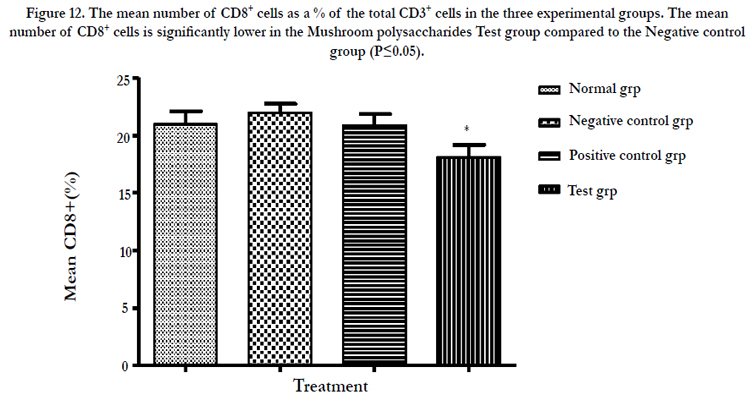
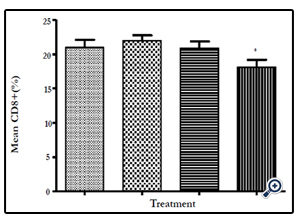
Figure 12: The mean number of CD8+ cells as a % of the total CD3+ cells in the three experimental groups. The mean number of CD8+ cells is significantly lower in the Mushroom polysaccharides Test group compared to the Negative control group (P≤0.05)
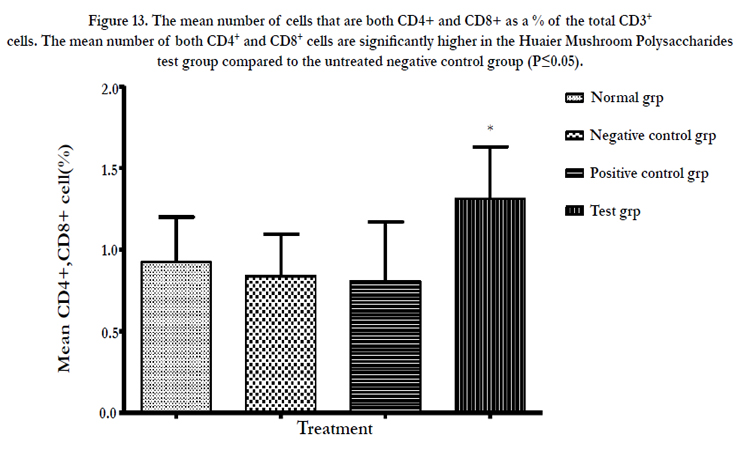
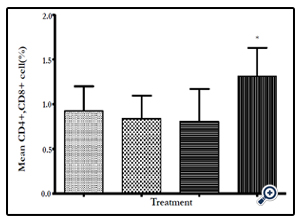
Figure 13: The mean number of cells that are both CD4+ and CD8+ as a % of the total CD3+ cells. The mean number of both CD4+ and CD8+ cells are significantly higher in the Huaier Mushroom Polysaccharides test group compared to the untreated negative control group (P≤0.05).
The Intestinal epithelial cells have been shown to express IL 7 mRNA in vitro elsewhere in this work. More so when the cells were subjected to polysaccharide extracts from fungi grown on media supplemented with the Chinese herb Gancao residues. In order to ascertain that this was also the case in vivo, a small section of mouse small intestine was excised from all four experimental groups and total RNA and cDNA obtained using conventional methods. Two genes GAPDH and IL 7 were amplified using quantitative real time PCR. Relative to GAPDH mRNA, the expression of IL 7 mRNA was 2.5 times higher in mice receiving the test drug (polysaccharide extracts) and was significantly different from the reference normal mice. There was no significant change in levels of expression of IL 7 mRNA in the negative control mice (Table 8, Figure 14).
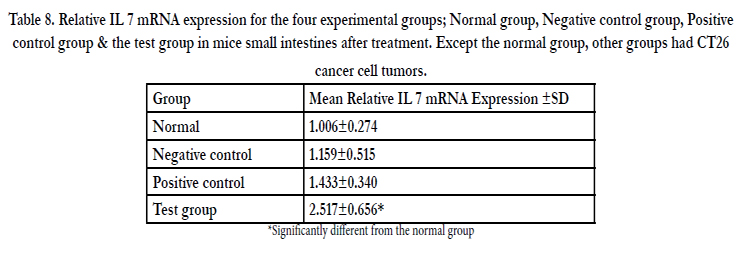
Table 8. Relative IL 7 mRNA expression for the four experimental groups; Normal group, Negative control group, Positive control group & the test group in mice small intestines after treatment. Except the normal group, other groups had CT26 cancer cell tumors.
*Significantly different from the normal group
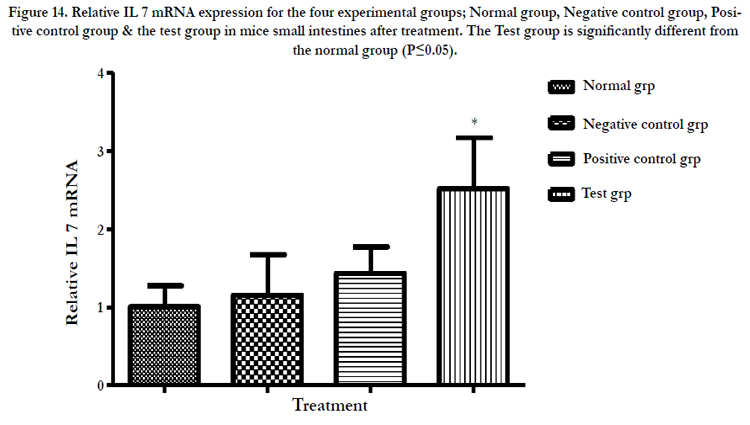
Figure 14. Relative IL 7 mRNA expression for the four experimental groups; Normal group, Negative control group, Positive control group & the test group in mice small intestines after treatment. The Test group is significantly different from the normal group (P≤0.05).
A standard curve was drawn and used to estimate the concentration of IL 7 in the samples of Normal group, Negative control group, Positive control group (Cytoxin group) and Test group (Receiving the polysacharride extract). OD value was located on the Y- axis and a horizontal line extended to the standard curve. At the point of intersection, a vertical line was drawn to the Xaxis and the corresponding concentration read. The concentrations in peripheral blood were determined to be approximately 15, 16, 18 & 19 pg/ml while in the intestinal mucosa 5, 12, 12.5 & 16 pg/ml for the normal, negative control, positive control and test groups respectively (Table 9, figure 15&16).
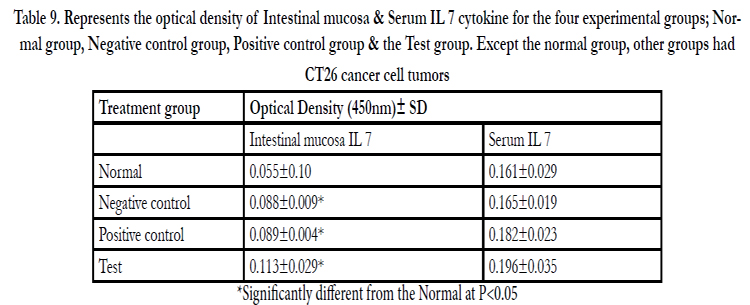
Table 9. Represents the optical density of Intestinal mucosa & Serum IL 7 cytokine for the four experimental groups; Normal group, Negative control group, Positive control group & the Test group. Except the normal group, other groups had CT26 cancer cell tumors.
*Significantly different from the Normal at P≤0.05
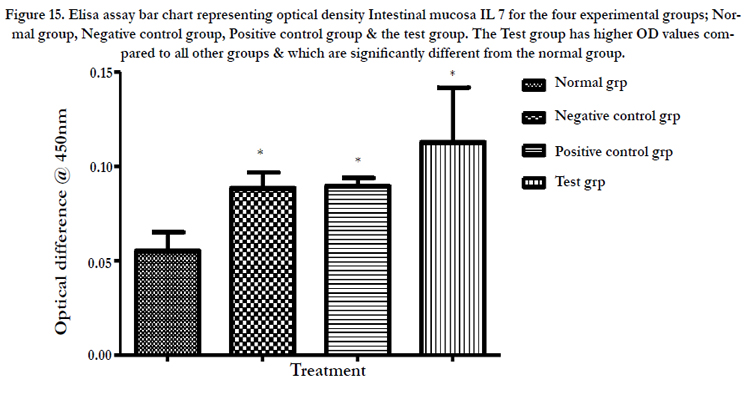
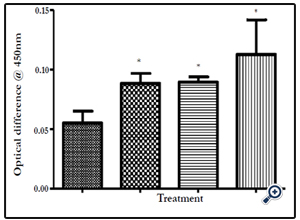
Figure 15. Elisa assay bar chart representing optical density Intestinal mucosa IL 7 for the four experimental groups; Normal group, Negative control group, Positive control group & the test group. The Test group has higher OD values compared to all other groups & which are significantly different from the normal group.
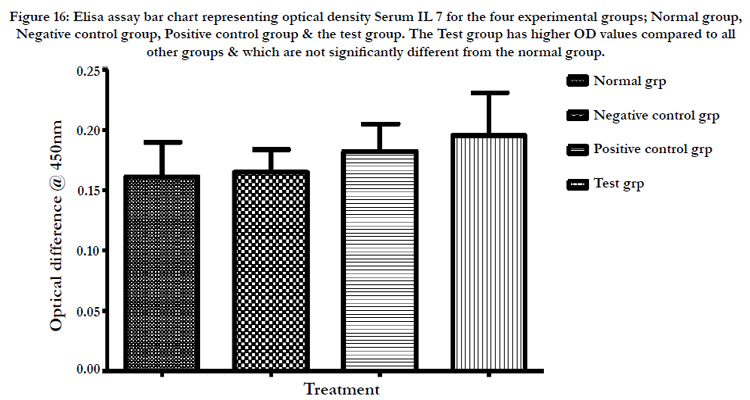
Figure 16: Elisa assay bar chart representing optical density Serum IL 7 for the four experimental groups; Normal group, Negative control group, Positive control group & the test group. The Test group has higher OD values compared to all other groups & which are not significantly different from the normal group.
Levels of Cytokines some of which share the common receptor gamma chain (γc), a component of their receptors and classically defined as growth and survival factors with IL 7 were also assayed in peripheral blood. They have been found to improve the survival and proliferation of T cells in vitro [31]. These included IL 2 and IL 15. IL 2 (19.807±10.959‡) & IL 15 (266.000±119.025*) levels were found to be significantly raised compared to either the normal animal group or negative control group. IL 6 and IL 10 involved in humoral immunity [32] were equally raised compared to either the normal animal group or negative control group. TNFα which supports cell-mediated immunity was found to be significantly. (P≤0.05) different from the normal group, higher levels being seen in polysaccharides treated group (16.042±1.158*), (Table 10, Figure 17 A, B, C, D, E & F).
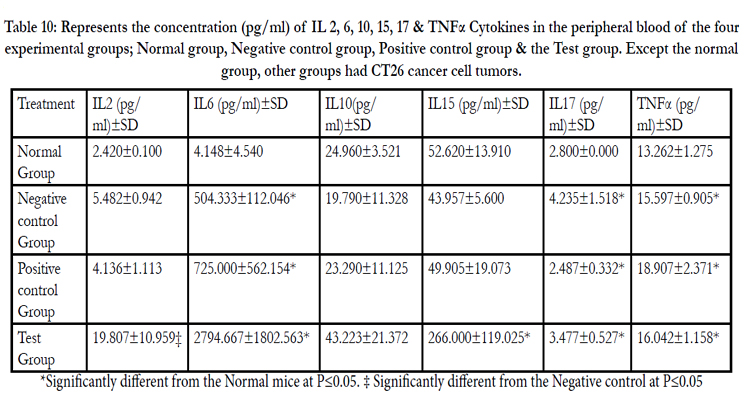
Table 10: Represents the concentration (pg/ml) of IL 2, 6, 10, 15, 17 & TNFα Cytokines in the peripheral blood of the four experimental groups; Normal group, Negative control group, Positive control group & the Test group. Except the normal group, other groups had CT26 cancer cell tumors.
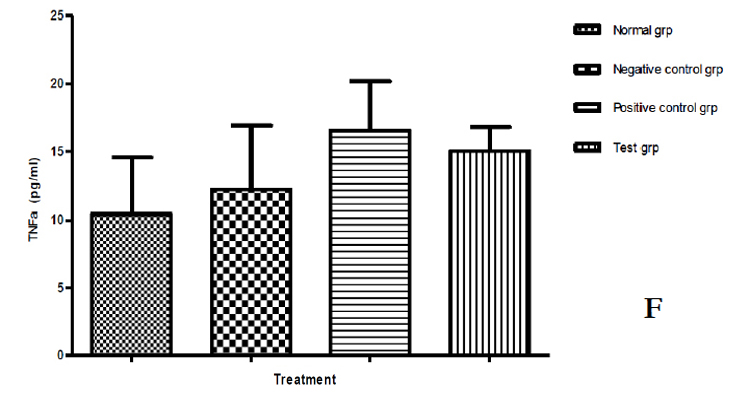
Figure 17: Levels of Cytokines which share the common receptor gamma chain (γc), with IL 7 in the four experimental groups. A = IL 2, B = IL 6, C = IL 10, D = IL 15, E = IL 17 & F = TNFα.
Discussion
From cell proliferation data, Trametes robiniophila murr (Huaier) polysaccharide extracts were observed to have minimal or no cytotoxicity to IEC-6 cells. Huaier polysaccharide extracts reportedly have no cytotoxicity to L-929 normal cells [33]. Mushrooms polysaccharides have been consumed as food stuff for centuries with over 3000 species reportedly being edible and 700 among them being medicinal with only 1% reported as poisonous [34]. Mushroom polysaccharide extracts from the Chinese mushroom Trametes robiniophila murr (Huaier) are therefore relatively safe for consumption as dietary supplements or Phytomedicines. IEC-6 cells exposed to mushroom polysaccharide extracts at two concentrations of 100µg/ml and 3.33mg/ml positively expressed IL-7 cytokine. Intestinal epithelial cells (IEC) have been implicated in IL-7 synthesis before [21]. IL-7 has been associated with potential for adoptive immunotherapy [13]. IL-7 cytokine produced can be associated with the immunomodulation medicinal value present in this species of mushroom. Mushrooms have been previously reported as having protective mechanism against cancer is their capacity to stimulate the immune system response where beta-glucan, a water-soluble polysaccharide is presumed to activate certain immune cells and proteins that attack cancer, including macrophages, T-cells, natural killer cells, and interleukins [5,35,36]. The cells exposed to 6 hours and 3 hours express IL-7 mRNA in a time dependent way. The expression at 3 hours was not significantly different from the control but a significant difference was noticed on exposure to 6 hours. At 100µg/ml the expression of IL7 is better than at 3.33mg/ml. The difference might be explained away as a result of shock due to sudden exposure to a high dose despite lack of noticeable toxicity therefore possibly needs more time as shown by results due to exposure for 72hours. Cells might thus require a longer adjustment time to the new micro-environment. In the lower dose cells continue with normal metabolic processes catalyzed by the extracts hence the better expression at the lower dose. The lower dose would possibly be more acceptable in drug dosage design.
The thymus weight indexes (figure 13) in the tumor cells infected test groups of mice are significantly lower than in the untreated healthy mice. This is possibly indicative of the role played by thymus gland in immune responses to tumor cells. The production of lymphocytes in great numbers possibly leading to eventual degeneration of the organ as happens in nature with age. The indices are also a reflection of higher mean weights in cancer cases when using the mushroom polysaccharides extract. This could be a positive indicator of the overall usefulness of Trametes robiniophila polysaccharide extracts when considered in totality as a nutritional form of treatment. Thymus gland undergoes slower atrophy in healthy mice as it is not subjected to undue pressure to raise the level of immunity. The function of thymus gland in immune responses is demonstrated in this research. The converse is true for the spleen weight indices. The spleen indices (figure 14) are significantly higher in mice infected with tumor cells and more so higher in mice treated with mushroom polysaccharides. This may be indicative of formation of more splenocytes as a response to the positive challenge by the tumor cells. This too may be indicative of better treatment outcome in mice subjected to the mushroom polysaccharides. This may be a further demonstration of the immunological response including expansion of the lymphocytes pool initiated in the spleen due to the presence of the foreign cancer cells. T-cells stored in the spleen are activated and mobilized to perform their immune functions.
The treatments affect Lymphopoiesis differently with the effect being spread across T lymphocytes CD4+ and CD8+ under consideration. In vivo, administration of IL-7 has been demonstrated to enhance peripheral T-cell functional capacity and expand the peripheral T-cell population [22]. Geiselhart et al. [37] have reported that IL-7 administration alters the peripheral Tcell CD4:CD8 ratio, and results in increases in peripheral T-cell numbers and altered function. The percentage of CD8+ T cells is higher in the spleen, leading to an inverse CD4/CD8 ratio. Thus, the distribution of the different lymphocyte subsets is markedly different between spleen and peripheral blood, inferring an important and distinct role for the spleen in CD4+ and CD8+ T cell activation [38].
There is a possibility that other organs besides the thymus and spleen are involved in generation and or maturation of T-lymphocytes. The Intestinal mucosa could possibly be a source of the T lymphocytes. The higher proportion of CD4+ and CD8+ cells (Figures 15, 16, & 17) in the Mushroom Polysaccharides treatment group comparable to conventional treatment is thought to be indicative of better prognosis in disease outcome. CD4+ T cells, also known as T-helper (Th) cells, play an important role in orchestrating adaptive immune responses to various infectious agents [27]. They are also involved in the induction of autoimmune and allergic diseases. Upon T-cell receptor (TCR)-mediated cell activation, naive CD4+ T cells can differentiate into at least four major lineages, Th1, Th2, Th17, and iTreg cells that participate in different types of immune responses [27]. This study has confirmed the postulation that IL-7 is thought to be produced by the intestinal epithelial cells (IEC) and, IL-7 has been indicated as having potential for adoptive immunotherapy and is capable in vivo of causing CD4+ T cell dependent destruction of tumor cells [13]. CD4+T cells play a critical role in the development of effective anti-tumor immunity [14]. IL-7 is thought to function in the immune system by providing the right cells in sufficient numbers to support and improve specific immune responses against infectious agents and malignant cells [29]. Mushroom polysaccharides would seem to rightly so as speculated to be acting as the stimulants in IEC IL -7 synthesis which leads to the cascade events leading to higher CD4+ cells observed. The mechanism of action of Mushroom polysaccharides could therefore be through stimulation of Intestinal Epithelial Cells which causes up regulation of IL -7 which then stimulates immune organs including the intestinal mucosa to produce more CD4+ and CD8+ lymphocytes as well as ensures their long survival. This is supported by the up regulation of IL 7 mRNA to 2.5 times in the small intestines in the test group. Positive effect of mushroom polysaccharides is also confirmed by the lower mortality rate (Table 5 and figure 12) in this group at the end of assay and lower tumor weight indices. Mushroom polysaccharides have been shown to produce over 50% reduction in tumor size and prolong the survival time of tumor bearing mice [5].
Elevated levels of IL-7 in serum could be associated with T-cell lymphopenia and, could be reflecting a homeostatic response, recovery of CD4+ counts should therefore lead to a decline in IL-7 levels. The levels of IL-7 observed in the circulation are likely to reflect more significant alterations of available IL-7 within the tissue microenvironment in the thymus and would be compatible with IL-7 as a primary regulator of T-cell homeostasis. Similarly, circulating IL-7 levels might only rise after increases have already occurred in the thymus tissue to a level sufficient to allow “spillover” into the circulation. Elevation of IL-7 within the lymphoid microenvironment in mice with CD4+ depletion contributes to the increased peripheral expansion, accumulation of activated cells, and thymic rebound observed CD4-depleted hosts [39,40].
The inducible IL-7 production and enhanced IL-7 activity after lymphopenia are implicated in the activation of CD4+ and CD8+ T cells in autoimmune diseases, such as rheumatoid arthritis, experimental autoimmune encephalomyelitis and type I diabetes [41]. Infiltration of the tumors by the CD4+ and CD8+ cells signifies better prognosis in cancer cases [18]. The contact circulating lymphocytes have with cancer cells is useful for Immunosurveillance because their deficit favors malignancy progression [37]. The polysaccharide extract from Trametes robiniophila is able also to stimulate increased levels of IL-6 and TNF-α which are associated with inflammatory effects linked to anti-tumor activity. IL-2 on the other hand is expressed by activated CD4+ & CD8+ T cells. It could therefore be confirming the role of Polysaccharide extracts tested in up regulating the expression of IL- 7 and production of IL-7 cytokine responsible for higher levels of activated CD4+ & CD8+ cells. IL-2 protects against autoimmunity [31,32]. Maintaining equilibrium between these two cytokines is thought to have therapeutic potential and Trametes robiniophila polysaccharide extracts have been demonstrated in the current research to have the potential to help maintain this balance. Botanical polysaccharides mushroom polysaccharides included, exhibit a number of beneficial therapeutic properties and the mechanisms involved in these effects could also be due to the modulation of innate immunity and, more specifically, macrophage function. The Huaier polysaccharide extract is considered in this study as being useful in the fight against Cancer.
Conclusion
Polysaccharide extracts obtained from mushroom Trametes robiniophila murr grown on media supplemented with Chinese herb Gancao (G.glabra L.) up regulates IL-7 mRNA both in vitro and in vivo in Intestinal epithelial cells. In vivo IL-7 cytokine transcription increases in the intestinal mucosa. CD4+ cells levels increase in the peripheral blood. Huaier polysaccharide extracts offer an alternative treatment to cancer cases (figure 18). It can be employed as a Nutraceutical for immunotherapy and Immunosurveillance. The mechanism of action has been deduced to probably be stimulation of IEC cells to produce increased IL-7 Cytokine responsible for resultant adaptive immunity and cancer immuno surveillance. The inclusion of Trametes robiniophila murr polysaccharide extracts as a Nutraceutical in the management of Cancer patients is recommended.
Figure 18. Mechanism of action of Huaier polysaccharides extracts
Acknowledgement
I wish to acknowledge the kind provision of a study scholarship by the China Scholarship Council (CSC) through International Education College of Tianjin University of Traditional Chinese Medicine and my employer the Kenya Medical Research Institute (KEMRI) for allowing me a three year study leave.
References
- Fact sheet N°297. October 2011. WHO.
- Tony S. Mok MD, Qing Zhou MD, Yi-Long Wu MD (2012) Research and Standard Care: Lung Cancer in China. American Society of Clinical Oncology 432-436.
- Margaret McDonald, Robin P. Hertz, Susan W. Pitman Lowenthal (2008) The burden of cancer in Asia. Pfizer facts, Pfizer Medical division, USA. 92.
- Zaidman BZ, M. Yassin, J. Mahajna, S.P. Wasser (2005) Medicinal mushroom modulators of molecular targets as cancer therapeutic. Appl. Microbiol. Biotechnol 6:453–468.
- Wasser SP (2002) Medicinal mushrooms as a source of anti-tumor and immunomodulating polysaccharides. Applied journal of Microbiology Biotechnology 60(3): 258–74.
- Brown GD, Herre J, Williams DL, Willment JA, et al. (2003) Dectin-1 mediates the biological effects of beta-glucans. Journal of Experimental Medicine 197:1119–1124.
- Wei S, Helsper JPFG, Van Griensven LJLD (2008) Phenolic compounds present in medicinal mushroom extracts generate reactive oxygen species in human cells in vitro. Int. Journal of Medicinal Mushrooms 10:1–13.
- Shamtsyan MM, Konusova VG, Goloshchev AM, Maksimova YO, Panchenko AV, et al. (2004) Immunomodulating and anti-tumor effects of basidiomycetes Pleurotus ostreatus (jacq.: fr.) P. Kumm. and P. cornucopiae (Pau.Ex Pers.) Rollan. Journal of Biological Physiological Chemistry 4(3):157 –61.
- Chen H.S, Tsai Y.F, Lin S, Lin CC, Khoo K.H (2004) Studies on the immuno-modulating and antitumor activities of Ganoderma Lucidum (Reishi) polysaccharides. Bioorganic and Medicinal Chemistry 12:5595– 5601.
- Chien C.M, Cheng J.L, Chang W.T, Tien M.H, Tsao C.M (2004) Polysaccharides of Ganoderma lucidum alter cell immunophenotypic expression and enhance CD56+ NK-cell cytotoxicity in cord blood. Bioorganic and Medicinal Chemistry 12:5603– 5609.
- Min B.S, Gao J.J, Nakamura N, Hattori M (2000) Triterpenes from the spores of Ganoderma lucidum and their cytotoxicity against Meth-A and LLC tumor cells. Chemical and Pharmaceutical Bulletin 48:1026–1033.
- Li L, Ye S, Wang Y, Tang Z (2006) Progress on experimental research and clinical application of Trametes robiniophila. China Cancer 16: 110–3.
- Itisam Sarangi, Dipanjan Ghosh , Sujit Kumar Bhutia, Sanjaya Kumar Mallick, Tapas K. Maiti (2006) Anti-tumor and immunomodulating effects of Pleurotus ostreatus mycelia-derived proteoglycans. International Immunopharmacology 6: 1287 – 1297.
- Feng CG, Zheng L, Jankovic D, Bafica A, Cannons JL,et al. (2008)The immunity-related GTPase Irgm1 promotes the expansion ofactivated CD4+ T cell populations by preventing interferon-gamma-induced cell death. Natural Immunology 9:1279-87.
- Kennedy R, Celis E (2008). Multiple roles for CD4+ T cells in anti-tumor immune responses. Immunological Reviews 222:129–144.
- Besser MJ, Shapira-Frommer R, Treves AJ, Zippel D, Itzhaki O, et al. (2010) Clinical responses in a phase II study using adoptive transfer of short-term cultured tumor infiltration lymphocytes in metastatic melanoma patients. Clinical Cancer Research 16:2646–2655.
- Dudley ME, John R. Wunderlich, James C. Yang, Richard M. Sherry, Suzanne L. Topalian, et al (2005) Adoptive cell transfer therapy following non-myeloablative but lymphodepleting chemotherapy for the treatment of patients with refractory metastatic melanoma. Journal of Clinical Oncology 23:2346–2357.
- Klebanoff C. A, Acquavella N, Yu Z, Restifo N. P (2011) Therapeutic cancer vaccines:are we there yet? Immunological Reviews 239: 27–44.
- Eiichi Sato, Sara H. Olson, Jiyoung Ahn, Brian Bundy, Hiroyoshi Nishikawa,Et al (2005) Intraepithelial CD8+ tumor-infiltrating lymphocytes and a high CD8+/regulatory T cell ratio are associated with favorable prognosis in ovarian cancer. Proc Natl Acad Sci U S A 102(51):18538-43.
- Marc Pellegrini, Thomas Calzascia, Alisha R Elford, Arda Shahinian, Amy E Lin, et al (2009) Adjuvant il-7 antagonizes multiple cellular and molecularinhibitory networks to enhance immunotherapies. Natural Medicine 15:528–536.
- Madrigal-Estebas L, McManus R, Byrne B. (1997) Human small intestinal epithelial cells secrete interleukin-7 and differentially express two different Interleukin-7 mRNA transcripts: implications for extrathymic T-cell differentiation. Hum Immunol. 58:83-90.
- Hua Yang, Ariel U. Spencer, Daniel H. Teitelbaum (2005) Interleukin-7 administration alters intestinal intraepithelial Lymphocyte phenotype and function in vivo. Cytokine 31: 419- 428.
- Akdis M. MD PhD, Simone Burgler PhD, Reto Crameri PhD, Thomas Eiwegger MD, Hiroyuki Fujita MD PhD, et al (2011) Interleukins, from 1 to 37, and interferon-g: Receptors,functions, and roles in diseases. Journal of allergy and clinical immunology 127(3):701-721.
- Capitini CM, Chisti AA, Mackall CL (2009) Modulating T-cell homeostasis with IL-7: preclinical and clinical studies. Journal of Internal Medicine 266:141-53.
- Palm NW, Medzhitov R (2009) Pattern recognition receptors and control of adaptive immunity. Immunological Reviews 227: 221–233.
- Paust S, Senman B,Von Andrian UH (2010) Adaptive immune responses mediated by natural killer cells. Immunological Reviews 235: 286–296.
- Rutishauser RL, Kaech SM (2010) Generating diversity: transcriptional regulation of effector and memory CD8+ T-cell differentiation. Immunological Reviews 235: 219–233.
- Zhu J, Paul WE (2010) Peripheral CD4+ T-cell differentiation regulated by networks of cytokines and transcription factors. Immunological Reviews 238: 247– 262.
- Cytheris - Interleukin-7. The immune enhancing company (2012).
- Mizuno T (1996) Development of antitumor polysaccharides from mushroom fungi. Food and Food Ingredient Japanese Journal 167: 69-87.
- Shoshana D. Katzman, Katrina K. Hoyer, Hans Dooms, Iris K. Gratz, Michael D. Rosenblum, et al. (2011) Opposing functions of IL-2 and IL-7 in the regulation of immune responses. Cytokine 56(1): 116–121.
- Murphy KM, Reiner SL (2002). The lineage decisions of helper T cells. Nat Rev Immunology 2:933–44.
- Yi Sun, Tiewei Sun, Feng Wang, Jing Zhang (2013) A polysaccharide from the fungi of Huaier exhibits anti-tumor potential and immunomodulatory effects. Carbohydr Polym 92 (1):577-82.
- Borchers AT, Stern JS, Hackman RM (1999) Mushrooms, tumors, and immunity. Proc Soc Exp Biol Med 221:281-293.
- Velvicka V, Vashishta A, Saraswat-Ohri S, Velvickova J (2008) Immunological effects of yeast and mushroom-derived beta glucans. Journal of Medicinal Food 11:615–22.
- Ikekawa T (2001). Beneficial effects of edible and medicinal mushrooms on health care. International Journal of Medicinal Mushrooms 3:291–8.
- Geiselhart LA, Humphries CA, Gregorio TA, Mou S, Subleski J, et al. (2001) IL-7 administration alters the CD4:CD8 ratio, increases T cell numbers, and increases T cell function in the absence of activation. Journal of Immunology 166: 3019-27.
- Nolte MA, Hamann A, Kraal G, Mebius RE (2002) The strict regulation of lymphocyte migration to splenic white pulp does not involve common homing receptors. Immunology 106:299–307.
- Terry J. Fry, Elizabeth Connick, Judith Falloon, Michael M. Lederman, David J. Liewehr,et al (2001). A potential role for interleukin-7 in T-cell homeostasis. Blood 97: 2983-2990.
- Terry J. Fry, Marcin Moniuszko, Stephen Creekmore, Susan J. Donohue, Daniel C. Douek, et al (2003). IL-7 therapy dramatically alters peripheral T-cell homeostasis in normal and SIV-infected nonhuman primates. Blood 101:2294-2299.
- Debaprasad Mandal, Lakshmishri Lahiry, Arindam Bhattacharyya, Sankar Bhattacharyya, Gaurisankar Sa, et al (2006) Tumor-induced thymic involution via inhibition of IL-7Ra and its JAK-STAT signaling pathway: Protection by black tea. International Immunopharmacology 6: 433– 444





