Mechanisms of Increased Survival in Lipopolysaccharide-Treated Mice With a Single Subcutaneous Capsaicin Challenge
Ijiri Y1*, Kato R1, Sasaki D1, Takano M1, Naruse M1, Hannya N1, Furukawa Y1, Inoue M1, Tomi R1, Hosako S1, Unno M1, Tanikawa S1, Tsukura Y1, Okada Y2, Amano F3, Matsuda N4, Tanaka K5, Hayashi T1
1 Cardiovascular Pharmacotherapy and Toxicology, Osaka University of Pharmaceutical Sciences, 4-20-1 Nasahara, Takatsuki, Osaka 569-1094, Japan.
2 Department of Pathology, Osaka Medical College, 2-7 Daigaku-machi, Takatsuki, Osaka 569-8686, Japan.
3 Biodefense and Regulation, Osaka University of Pharmaceutical Sciences, 4-20-1 Nasahara, Takatsuki, Osaka 569-1094, Japan.
4 Department of Emergency and Critical Care Medicine, Nagoya University Graduate School of Medicine, 65 Tsurumai-cho, Showa-ku, Nagoya, Aichi 466-0065, Japan
5 Shirasagi Hospital, 7-11-23 Kumata, Higashisumiyosi-ku, Osaka 546-0002, Japan.
*Corresponding Author
Yoshio Ijiri
Cardiovascular Pharmacotherapy and Toxicology, Osaka University of Pharmaceutical Sciences, 4-20-1 Nasahara,
Takatsuki, Osaka 569-1094, Japan.
Tel/Fax: +81-72-690-1045
E-mail: ijiri@gly.oups.ac.jp
Received: February 26, 2016; Accepted: April 14, 2016; Published: April 22, 2016
Citation: Ijiri Y, et al. (2016) Mechanisms of Increased Survival in Lipopolysaccharide-Treated Mice With a Single Subcutaneous Capsaicin Challenge. Int J Clin Pharmacol Toxicol. 5(2), 202-20 8. DOI : dx.doi.org/10.19070/2167-910X-1600034
Copyright: Ijiri Y© 2016. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
Background:Capsaicin, a pungent compound present in capsicum fruits, has various biological activities such as stimulation of transient receptor potential vanilloid-1 (TRPV-1) and/or inhibition of tumor necrosis factor-alpha (TNF-α). This study aimed to reveal the mechanism involved in capsaicin-mediated high survival rate in LPS-treated mice.
Methods:In LPS-treated mice, a comparative survival rate study using a TRPV-1 agonist (capsaicin) and a TRPV-1 antagonist (capsazepine). DNA-binding activity of NF-κB and TUNEL staining was conducted in vivo. In LPS-treated RAW264 cells, TNF-α was measured by enzyme-linked immunosorbent assay.
Results: In the present study, the survival rate of mice treated with 20 mg/kg LPS was significantly improved from 7.7% to 92.3% after treatment with 4 mg/kg capsaicin and to 53.8% with 40 mg/kg capsazepine. In liver tissues, the LPS-induced increase in NF-κB DNA-binding activity was attenuated by capsaicin. The increased number of TUNEL-positive cells observed in the alveolar epithelial and liver parenchymal cells of LPS-treated mice was diminished by capsaicin treatment. In RAW264 cells, mouse leukemic monocytes, capsaicin inhibited the release of TNF-α after LPS stimulation, by attenuating NF-κB DNA-binding activity.
Conclusion: The improvement in the survival rate (92.3% -high) by capsaicin administration in LPS challenge could be explained not only by TRPV-1 activity, but also by the pleiotropic effect which inhibits TNF-α release from macrophages. Thus, capsaicin may be a promising treatment option for LPS-related diseases such as severe sepsis or septic shock.
2.Abbreviations
3.Introduction
4.Materials and Methods
4.1.Reagents
4.2.Animals
4.3.Treatment of mice
4.4.Survival rate experiments
4.5.Preparation of embedded samples
4.6.HE and TUNEL stains
4.7.Estimation of NF-κB DNA-binding activity
4.8.Cell culture
5.Determination of Tnf- a Concentration in Raw264 Cells Using Enzyme-Linked Immunosorbent Assay (ELISA)
5.1.Statistical Analysis
6.Results
6.1.Survival tests
6.2.Histological examination
6.3.NF-κB DNA-binding activity
6.4.TNF-α release from mouse leukemic monocytes, RAW264 cells
7.Discussion
8.Acknowledgement
9.References
Keywords
Capsaicin, Capsazepine, Lipopolysaccharide, Nuclear Factor-Kappa.
Abbreviations
DAB-Diaminobenzidine; DTT-dithiothreitol; IL-interleukin; HE-Hematoxylin-Eosin, LPS-lipopolysaccharide;
NF-κB-nuclear factor kappa-light-chain-enhancer of activated B; NO-nitric oxide; PBS-phosphate buffered saline;
PPAR& gamma;-peroxidase proliferater-activated receptor gamma; TDT-terminal deoxynucleotidyl transferase; TLR-toll-like receptor; TNF-α-tumor necrosis factor-alpha; TNF-R-tumor necrosis factor-receptor; TRPV1-transient receptor potential vanilloid 1; TUNEL-terminal deoxynucleotidyl transferase dUTP nick end labeling.
Introduction
Absolute mortality in severe sepsis has been decreased from 35.0% to 18.4% [1]. However, when if early aggressive treatment is not started (example; after suspected less than 6 h), the outcome is usually poor. Severe sepsis and septic shock are thought to result from the overproduction of cytokines (proinflammatory mediators), including tumor necrosis factor-alpha (TNF-α), interleukins (IL-1β, IL-6, IL-10) [2,3], and nitric oxide (NO) [4, 5, 6, 7]. However, the disease etiology is completely unknown. Recently, a preliminary report investigated the clinical efficacy of continuous hemodiafiltration (CHDF) with an AN69ST hemofilter (AN69ST-CHDF) for patients with septic shock [8], but studies with TNF-α removal medications are not yet conducted in the clinical setting. The survival rate after lipopolysaccharide (LPS)-induced endotoxic shock in mice was reported as 73.3% with cerivastatin treatment [6] and 60% with virgin olive oil treatment [7]. Furthermore, we have previously reported that capsaicin (4 mg/kg, subcutaneously; s.c.) inhibits the overexpression of TNF-α, IL-1β, and NOx levels and has the potential to improve survival rate (from 0% to 91.7%) in LPS (20 mg/kg, intraperitoneally; i.p.) treated mice [4].
Capsaicin (trans-8-methyl-N-vanillyl-6-nonenamide), a pungent compound present in capsicum fruits, is synthesized by the addition of a branched-chain fatty acid to vanillylamine.
Cap saicin and several related compounds are called capsaicinoids and are produced as secondary metabolites by chili peppers. Pure capsaicin is a volatile, hydrophobic, colorless, odorless, and crystalline to waxy compound. Capsaicin has various biological activities including transient receptor potential vanilloid-1 (TRPV- 1) activation [9, 10], and antioxidant [11, 12], antibacterial [13], and anti-inflammatory effects [14, 15]. It is used as an ingredient in spicy foods and in oral dietary supplements, as an analgesic in topical ointments, nasal sprays, and dermal patches. It is typically used to treat post-herpetic neuralgia, painful diabetic neuropathy, pain from osteoarthritis [16], pruritus [17], and in pepper spray aerosols [18] in clinical settings.
Capsaicin has a reported additive effect on the inhibition of gastric acid secretion, through activation of TRPV-1 in vivo [19]. Capsaicin has also been reported to suppress TNF-α production by LPS-stimulated macrophages in vitro by downregulating nuclear factor kappa-light-chain-enhancer of activated B (NF- κB) through peroxisome proliferator-activated receptor gamma (PPARγ) ligand-like activity [11, 20]. With respect to drug interactions, capsaicin upon large and long-term administration decreases the absorption of digoxin by inducing CYP3A2 mRNA expression via indirect activation of PXR in LPS-treated rats [21]. Moreover, pre-treatment of rats with capsaicin resulted in decreased bioavailability of simvastatin and galantamine [22, 23]. Additionally, other studies indicate that capsaicin at noncytotoxic doses (less than 100 µM), causes functional and structural changes in Caco-2 cell monolayers [24].
To elucidate the relation between an improved survival rate after LPS challenge due to capsaicin administration and TRPV- 1 activation, a comparative survival rate study of the TRPV-1 agonist, capsaicin and the TRPV-1 antagonist, capsazepine (CPZ) [25, 26] was conducted. Furthermore, to clarify the interference between capsaicin and NF-κB, the DNA-binding activity of NF- κB was measured. Morphological changes were observed using HE staining under the light microscope upon LPS challenge in vivo (mice) and TNF-α levels were determined in vitro (RAW264 cells).
Materials and Methods
LPS (Escherichia coli, 055:B55, Lot No. 114K4107) and CPZ (TRPV-1 antagonist) were purchased from Sigma-Aldrich, and capsaicin (98% pure) was provided by Maruishi Pharmaceutical Co., Ltd. Sevoflurane was purchased from Maruishi Pharmaceutical Co., Ltd. The other reagents used were commercially available ultrapure grade chemicals.
Male BALB/c mice (aged 8- to 10-week-old; weight, 21-26 g; Japan SLC, Inc.) were used for these studies. They were acclimatized for at least one week to controlled environmental conditions (temperature, 24°C ± 1°C; humidity, 55% ± 10%; light cycle, 6:00-18:00 h) with free access to solid food (NMF, Oriental yeast Co., Ltd.) and water. All experimental procedures were approved according to the guidelines for the use of experimental animals, and were conducted in the animal facility established by the ethics committee of Osaka University of Pharmaceutical Sciences (approved on 17, March 2009, No. 09014). At the end of the experiment, mice were euthanized with sevoflurane.
LPS was dissolved to a concentration of 1 mg/mL in physiological saline (Fuso Pharmaceutical Industries Ltd.). Capsaicin and CPZ were dissolved in physiological saline with 1% ethanol and 1% Tween-20. LPS (20 mg/kg) was administered i.p., and capsaicin (4 mg/kg), and CPZ (40 mg/kg) were administered s.c. in the dorsal region of mice 5 min after LPS administration.
Mice were divided into the following eight treatment groups for the survival rate experiments: vehicle (physiological saline, i.p. + physiological saline, s.c., n = 13), LPS (LPS, i.p. + physiological saline, s.c., n = 13), capsaicin (physiological saline, i.p. + capsaicin, s.c., n=13), CPZ (physiological saline, i.p. + CPZ, s.c., n = 13), capsaicin + CPZ (physiological saline, i.p. + capsaicin and CPZ, s.c., n = 13), LPS + capsaicin (LPS, i.p. + Capsaicin, s.c., n=13), LPS + CPZ (LPS, i.p. + CPZ, s.c., n = 13), and LPS + capsaicin + CPZ (LPS, i.p. + capsaicin and CPZ, s.c., n = 13). Mice were monitored for mortality every 24 h for 7 days.
Each mouse was anesthetized with sevoflurane and restrained in a supine position. The heart was exteriorized and the left ventricle was cannulated for perfusion fixation. Physiological saline (20 mL) was perfused through the cannula, followed by 20 mL of 10% buffered formalin solution (Wako Pure Chemical Industries, Ltd.). After the perfusion fixation, the lungs and liver were removed. For paraffin embedding, fixation was performed for more than 24 h in 10% buffered formalin and tissues were then embedded in paraffin blocks for sectioning. The paraffin blocks were cut into 5-µm sections with a rotary microtome (Leica RM2125RT; Leica Microsystems Co., Ltd.). For frozen embedding, fixation was performed for more than 12 h in 30% sucrose and tissues were embedded in the Optimal Cutting Temperature compound (Sakura Finetechnical Co. Ltd.) for sectioning. The frozen embedded samples were cut into 7-µm sections with a cryotome (Leica CM1850, Leica Microsystems Co., Ltd.).
Paraffin sections were washed with tap water for 1.5 hours, dehydrated using standard methods, and stained with Mayer's H & E.
TUNEL stain was performed using an Apoptosis in situ Detection Kit (Wako Pure Chemical Industries, Ltd., Osaka, Japan), and stained tissues were observed and photographed with a light microscope (BX51; Olympus Co., Ltd.)
Blood was collected from the axillary artery of anesthetized mice at 0.5, 1, 3, and 6 h after administration of physiological saline or LPS. After ligating the portal vein and hepatic artery, the hepatic vein was cut, and the portal vein was perfused with physiological saline to remove the blood from the liver. One-hundred milligrams of hepatic tissue was lysed in 1 mL of lysis buffer [10 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) (pH 7.9), 1.5 mM MgCl2, 10 mM KCl, 1% dithiothreitol (DTT), 1% Protease Inhibitor Cocktail Set III, ethylenediaminetetraacetic acid (EDTA)-Free (Calbiochem)] before being homogenized with a homogenizer pestle. The homogenate was then centrifuged (20 min, 10000×g, 4°C) and the resulting precipitate was dissolved in 140 µL of extraction buffer [20 mM HEPES (pH 7.9), 1.5 mM MgCl2, 0.42 M NaCl, 0.2 mM EDTA, 25% glycerol, 1% DTT, 1% Protease Inhibitor Cocktail Set III, EDTA-Free] and incubated on ice (30 min). The suspension was then centrifuged (5 min, 18000×g, 4°C), and the supernatant was collected as nucleoprotein. DNA-binding activity of NF-κB in the liver tissues was measured using an NF-κB (p65) Transcription Factor Assay Kit (Cayman Chemical Company). The reaction was stopped by adding 100 µL/well of stop solution with gentle tapping. Within 5 min, absorbance at 450 nm was measured using a plate reader (Labsystems Multiscan MS; Dainippon Sumitomo Pharma Co., Ltd.). Data are represented as relative values with the assumption that DNA-binding activity of NF-κB in the vehicle group was 100%.
Mouse leukemic monocytes, RAW264 cells, were cultured in Dulbecco's modified Eagle's medium (DMEM, 4.5 g glucose/L, Nacalai Tesque, Inc.), containing 10% fetal bovine serum (BioSource International Inc.), penicillin (50 IU/mL), and streptomycin (50 IU/mL) (PS, Life Technologies Corporation) at 37°C with 5% CO2 in 25-cm2 tissue culture flasks (Becton-Dickinson Ind.). The RAW264 cells were sub-cultured by treatment with 0.25% trypsin and (1 mM) ethylenediaminetetraacetic acid (EDTA) in a Ca2+- and Mg2+-free PBS solution after every 3 days. This involved detachment of cells from the stock cultures by trypsin digestion, washing once by centrifugation, re-suspension, and sub-culture in 10 mL medium in culture flasks at a concentration of 5 × 106 cells/mL. The cultures were regularly observed under an inverted light microscope to monitor growth and contamination.
Determination of TNF-α Concentration in RAW264 Cells Using Enzyme-Linked Immunosorbent Assay (ELISA)
RAW264 cells (5 × 105 cells) were seeded on a 24-well plate and cultured for 3 days at 37°C with 5% CO2. The medium was aspirated and washed with PBS. After washing, medium containing LPS (100 ng/mL) and capsaicin (100 µM) was added to the cells and incubated for 1 h at 37°C with 5% CO2. After incubation, the medium from each well was collected and TNF-α concentration was determined by ELISA (Quantikine Immunoassay mouse TNF-α, R& D).
The survival rate data were plotted as Kaplan-Meier curves and examined for significant differences between the various treatment groups using the log-rank test. Results are shown as mean ± standard deviation (SD). Statistical analysis was performed using Tukey’s test. A significant difference was defined as p< 0.05.
Results
The comparative survival rate test at 7 days after the LPS challenge, and capsaicin (TRPV-1 agonist), and CPZ (TRPV-1 antagonist) treatment, was performed with 8 groups: vehicle, capsaicin, CPZ, capsaicin + CPZ, and LPS + capsaicin + CPZ groups (100%; 0 death of 13 participants); LPS + capsaicin group (92.3%; One death of 13 participants), LPS + CPZ group (53.8%; 6 deaths of 13 participants), and LPS group (7.7%; 12 deaths of 13 participants), where the mice died within 3 days. The vehicle, capsaicin, CPZ, capsaicin + CPZ, and LPS + capsaicin + CPZ groups (survival rate of 100%, respectively) and the LPS + capsaicin group (survival rate of 92.3%) showed a significantly improved survival rate for LPS (survival rate: 7.7% vs. 100% and 92.3%, (p < 0.01, respectively) and LPS + CPZ (survival rate: 53.8% vs. 100%, (p < 0.01 and survival rate: 92.3%,( p < 0.05, respectively) groups (Figure 1).
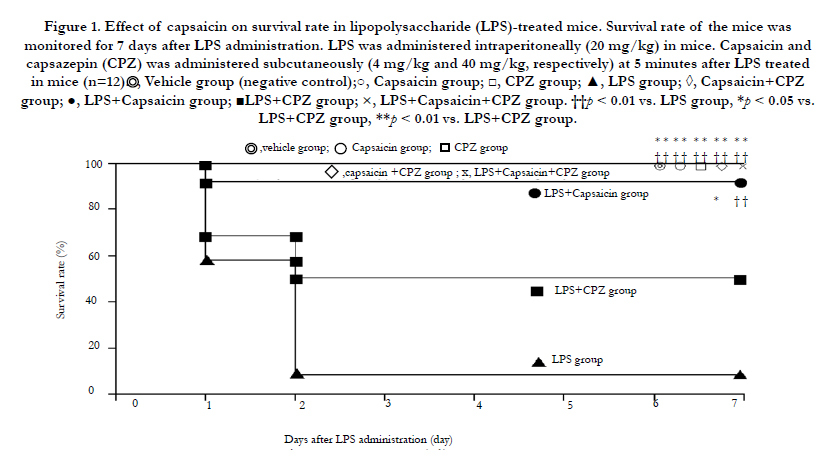
Figure 1. Effect of capsaicin on survival rate in lipopolysaccharide (LPS)-treated mice.
Histological examination of the HE-stained sections of lung tissues (terminal bronchioles and alveolar epithelial cells) removed at 12 h after LPS challenge, showed increased numbers of neutrophilic leucocytes (stab cells and segmented leucocytes) (Figure 2A). In the liver tissues at 12 h after LPS challenge, increase in spread sinusoids and diffused neutrophilic leucocytes (stab cells and segmented leucocytes) was observed (Figure 2B). On the other hand, 12 h after LPS + capsaicin treatment spread sinusoids and oil droplets were observed in the liver sections (Figure 2B). TUNEL-positive cells (TPCs) are indicated by brownish spots (Figure 3). The lung and liver tissues of the control and LPS + capsaicin-treated mice lacked TPCs, which were observed in the alveolar epithelial cells and liver parenchymal cells of the LPS group (Figure 3).
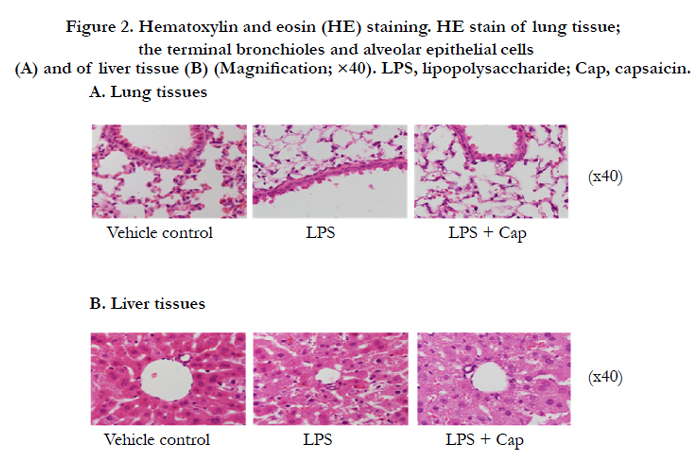
Figure 2. Hematoxylin and eosin (HE) staining. HE stain of lung tissue; the terminal bronchioles and alveolar epithelial cells (A) and of liver tissue (B) (Magnification; ×40). LPS, lipopolysaccharide; Cap, capsaicin..
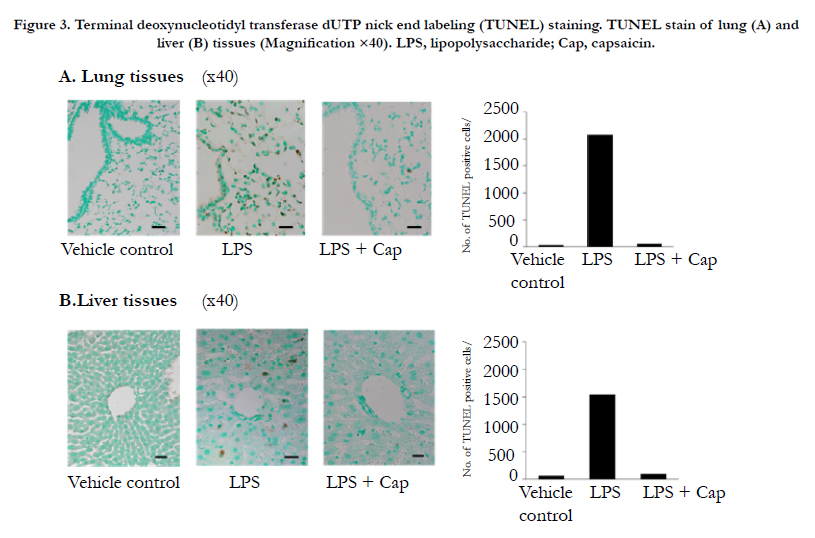
Figure 3. Terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) staining. TUNEL stain of lung (A) and liver (B) tissues (Magnification ×40). LPS, lipopolysaccharide; Cap, capsaicin..
Compared with the vehicle group, both the LPS and LPS + capsaicin groups showed significantly higher NF-κB DNA-binding activity at 1 h after LPS challenge ((p < 0.01, Figure 4). However, compared to the LPS group, the LPS + capsaicin group showed a significantly lower NF-κB DNA-binding activity at 1 h after LPS challenge ((p < 0.05, Figure 4).
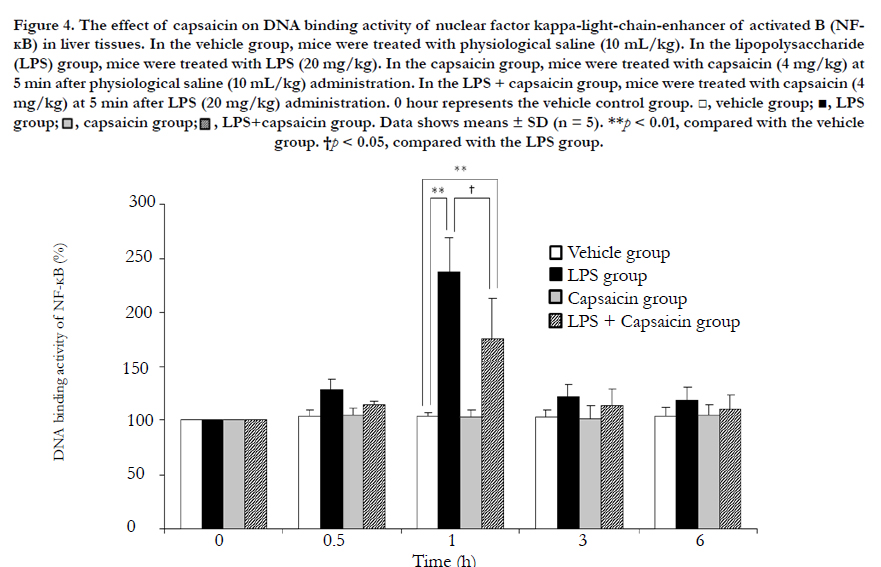
Figure 4. The effect of capsaicin on DNA binding activity of nuclear factor kappa-light-chain-enhancer of activated B (NFΚB) in liver tissues..
TNF-α release was measured after 1 h of LPS or capsaicin treatment using ELISA in RAW264 cells that are mouse leukemic monocytes. TNF-α release decreased significantly ((p < 0.05) in the capsaicin group, whereas the LPS and LPS + capsaicin groups showed significantly increased levels of TNF-α ((p < 0.01), compared to the control group. However, LPS + capsaicin treatment showed significantly decreased release of TNF-α compared to LPS treatment alone (p < 0.05, Figure 5).
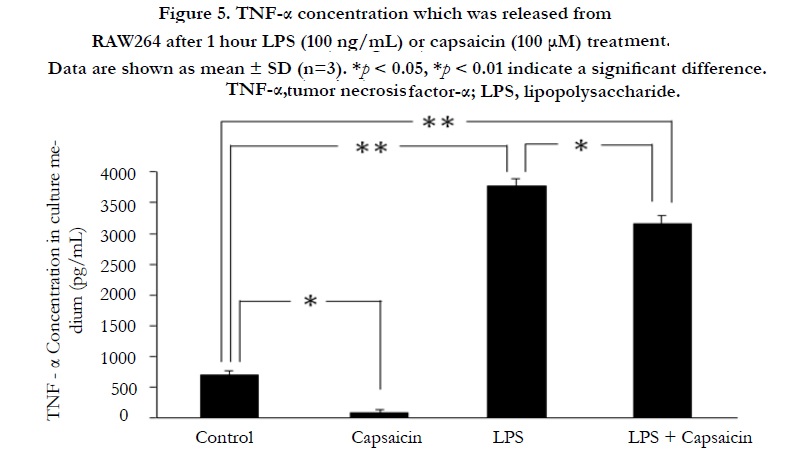
Figure 5. TNF-α concentration which was released from RAW264 after 1 hour LPS (100 ng/mL) or capsaicin (100 μM) treatment..
Discussion
Capsaicin has various biological activities including TRPV-1 activation [9, 10] and is used as an analgesic in topical ointments and dermal patches, typically to treat post-herpetic neuralgia, painful diabetic neuropathy, and osteoarthritic pain in the clinical setting [16] . Capsaicin is reported to have an additive effect on the inhibition of gastric acid secretion, through the activation of TRPV-1 (in vivo [19]. However, most of these clinical trials of capsaicin have been called off at phase 3. We have previously performed a dose-response trial of LPS (10, 15, and 20 mg/kg, i.p.) and capsaicin (2, 3, 4, 5, 10 mg/kg, s.c.); capsaicin (4 mg/kg, s.c.) significantly improved the survival (92.3%) of mice treated with LPS (20 mg/kg, i.p.)[4]. Furthermore, we also reported that capsaicin inhibited both plasma-soluble TNF production and mRNA expression of TNF-α [27]. Macrophages can also release TNF-α in the soluble form (sTNF) [28] that elicits biological activities by binding to type 1 and 2 TNF receptors (TNF-R1 and TNFR2) [29, 30, 31]. In this study, to investigate whether the improving action on survival rate by capsaicin in LPS challenge is via TRPV-1 or not, a comparative test of capsaicin (TRPV-1 agonist) vs. CPZ (TRPV-1 antagonist) was performed. In the LPS group, mice died within 3 days in the present study (survival rate: 7.7%, 12 death of 13 participants). In contrast, the survival rate in the LPS + capsaicin group (92.3%) was significantly improved, compared to that in the LPS + CPZ group. However, survival rate in the LPS + capsaicin + CPZ group was 100% and that in the LPS + CPZ group was 53.8%, which could not be explained by TRPV-1 action alone. The 53.8% survival rate with TRPV-1 antagonist treatment might be due to the pleiotropic effect of CPZ rather than TRPV-1 inhibition, while capsaicin has been reported to suppress TNF-α production in LPS-stimulated macrophages( in vitro by downregulating NF-κB via PPAR γ ligand-like activity [14, 15]. Furthermore, we also reported that capsaicin reduced circulating sTNF levels in LPS-treated mice by inhibiting the release of membrane-bound TNF and the expression of TNF-α mRNA [27]. In this study, the DNA-binding activity of NF-κB in the LPS + capsaicin group at 1 h after LPS challenge was significantly lower in the capsaicin treated group than that in the LPS group. Capsaicin prevented the nuclear translocation of NF-κB in liver tissues of LPS-treated mice and inhibited the appearance of TPCs in alveolar epithelial cells and liver parenchymal cells. In addition, capsaicin inhibited TNF-α release upon LPS challenge in RAW264 cells similar to Kupffer cells or alveolar macrophages [15]. These findings are consistent with reports that capsaicin inhibits TNF-α release from Kupffer cells or alveolar macrophages and inhibits TNF-α mRNA expression via modulation of NF-κB in LPS-treated mice [14, 15, 16, 17]. In HE staining, neutrophilic leucocytes as stab cells and segmented leucocytes were observed in lung tissues (terminal bronchioles and alveolar epithelial cells) and liver tissues at 12 h after the LPS challenge (Figure 2). In contrast, these cells were not observed in the vehicle and the LPS + capsaicin group. Capsaicin administration was found to cause oil droplets in liver parenchymal cells. Neutrophilic leucocytes were diffused to the alveolar and liver parenchymal cells after 12 h of LPS challenge. Thus, capsaicin might prevent the diffusion of neutrophilic cells. TPCs were highly enriched in liver parenchymal cells and alveolar epithelial cells at 12 h after the LPS challenge, compared with the vehicle and the LPS + capsaicin group, indicating that capsaicin treatment inhibited the expression of TPCs post LPS challenge. The decreased TPCs in alveolar epithelial cells and liver parenchymal cells might indicate that capsaicin prevented TNF-α production stimulated in Kupffer cells or alveolar macrophages by LPS challenge.
Our results support the mechanism that capsaicin might interfere with TNF-α mRNA transcription by inhibiting NF-κB DNA-binding activity via NF-κB suppression. Capsaicin-induced TNF-α mRNA suppression in alveolar tissues may be responsible for the survival benefit observed following the LPS challenge in this study.
Finally, the improving effect of capsaicin, resulting in 92.3% highsurvival rate upon LPS challenge can be explained not only by TRPV-1 action, but also by inhibition of TNF-α release from macrophages after the LPS challenge. Thus, capsaicin may be a promising treatment option for LPS-related diseases such as severe sepsis or septic shock.
Acknowledgement
The authors thank Dr. Akashi for the kind gift of RAW264 cells, and the Maruishi Pharmaceutical Co., Ltd. for providing Capsaicin.
References
- Kaukonen KM, Bailey M, Suzuki S, Pilcher D, Bellomo R (2014) Mortality Related to Severe Sepsis and Septic Shock Among Critically ill Patients in Australia and New Zealand, 2000-2012. JAMA 311(13): 1308-1316.
- Jayaraman P, Sada-Ovalle I, Nishimura T, Anderson AC, Kuchroo VK, et al. (2013) IL-1β promotes antimicrobial immunity in macrophages by regulating TNFR signaling and caspase-3 activation. J Immunol 190(8): 4196-4204.
- Xu W, Roos A, Schlagwein N, Woltman AM, Daha MR, et al. (2006) IL- 10-producing macrophages preferentially clear early apoptotic cells. Blood 107(12): 4930-4937.
- Tsukura Y, Takabatake Y, Nakazato H, Kato R, Ijiri Y, et al. (2007) Capsaicin Improves Survival Rate in Mice with Lipopolysaccharide-Induced Endotoxin Shock. Circ Cont 28(4): 307-313.
- MacMicking J, Xie QW, Nathan C (1997) Nitric oxide and macrophage function. Annu Rev Immunol 15: 323-350.
- Ando H, Takamura T, Ota T, Nagai Y, Kobayashi K (2000) Cerivastatin improves survival of mice with lipopolysaccharide-induced sepsis. J Pharmacol Exp Ther 294(3): 1043-1046.
- Leite MS, Pacheco P, Gomes RN, Guedes AT, Castro-Faria-Neto HC, et al. (2005) Mechanisms of increased survival after lipopolysaccharide-induced endotoxic shock in mice consuming olive oil-enriched diet. Shock 23(2):173-178.
- Shiga H, Hirasawa H, Nishida O, Oda S, Nakamura M, et al. (2014) Continuous hemodiafiltration with a cytokine-adsorbing hemofilter in patients with septic shock: a preliminary report. Blood Purif 38(3-4): 211-218.
- Binshtok AM, Bean BP, Woolf CJ (2007) Inhibition of nociceptors by TRPV1-mediated entry of impairment sodium channel blockers. Nature 449(7162): 607-610.
- Cortright DN, Szallasi A (2004) Biochemical pharmacology of the vanilloid receptor TRPV1. An update. Eur J Biochem 271(10): 1814-1819.
- Chen CW, Lee ST, Wu WT, Fu WM, Ho FM, et al. (2003) Signal transduction for inhibition of inducible nitric oxide synthase and cyclooxygenase-2 induction by Capsaicin and related analogs in macrophages. Br J Pharmacol 140(6): 1077-1087.
- Shimeda Y, Hirotani Y, Akimoto Y, Shindou K, Ijiri Y, et al. (2005) Protective Effects of Capsaicin against Cisplatin-Induced Nephrotoxicity in Rats. Biol Pharm Bull 28(9): 1635-1638.
- Kalia NP, Mahajan P, Mehra R, Nargotra A, Sharma JP, et al. (2012) Capsaicin, a novel inhibitor of the NorA efflux pump, reduces the intracellular invasion of Staphylococcus aureus. J Antimicrob Chemother 67(10): 2401-2408.
- Demirbilek S, Ersoy MO, Demirbilek S, Karaman A, Gürbüz N, et al. (2004) Small-dose Capsaicin reduces systemic inflammatory responses in septic rats. Anesth Analg 99(5): 1501-1507.
- Kim CS, Kawada T, Kim BS, Han IS, Choe SY, et al. (2003) Capsaicin exhibits anti-inflammatory property by inhibiting IkB-a degradation in LPSstimulated peritoneal macrophages. Cell Signal 15(3): 299-306.
- Anand P, Bley K (2011) Topical capsaicin for pain management: therapeutic potential and mechanisms of action of the new high-concentration capsaicin 8% patch. Br J Anaesth 107(4): 490-502.
- Gooding SMD, Canter PH, Coelho HF, Boddy K, Ernst E (2010) Systematic review of topical capsaicin in the treatment of pruritus. Int J Dermatol 49(8): 858-865.
- Reilly CA, Crouch DJ, Yost GS (2001) Quantitative analysis of capsaicinoids in fresh peppers, oleoresin capsicum and pepper spray products. J Forensic Sci 46(3): 502-509.
- Okumi H, Tashima K, Matsumoto K, Namiki T, Terasawa K, et al. (2012) Dietary agonists of TRPV1 Inhibit gastric acid secretion in mice. Planta Med 78(17): 1801-1806.
- Park JY, Kawada T, Han IS, Kim BS, Goto T, et al. (2004) Capsaicin inhibits the production of tumor necrosis factor alpha by LPS-stimulated murine macrophages, RAW 264.7: a PPARγ ligand-like action as a novel mechanism. FEBS Lett 572(1-3): 266-270.
- Kato R, Higashitani A, Irine J, Kusukawa Y, Yamamoto y, et al. (2012), Influence of capsaicin on fluctuation of digoxin pharama cokinetics in lippotysaccharide- treated rats.Xenobiotica 42: 798-807.
- Zhai XJ, Lu YN (2012) Food-drug interactions: effect of capsaicin on the pharmacokinetics of galantamine in rats. Xenobiotica 42(11): 1151-1155.
- Zhai XJ, Chen JG, Liu JM, Shi F, Lu YN (2013) Food-drug interactions: effect of capsaicin on the pharmacokinetics of simvastatin and its active metabolite in rats. Food Chem Toxicol 53: 168-173.
- Tsukura Y, Mori M, Hirotani Y, Ikeda K, Amano F, et al. (2007) Effects of capsaicin on cellular damage and monolayer permeability in human intestinal Caco-2 cells. Biol Pharm Bull 30(10): 1982-1986.
- Luo YL, Guo HM, Zhang YL, Chen PX, Zhu YX, et al. (2013) Cellular mechanism underlying formaldehyde-stimulated Cl- secretion in rat airway epithelium. PLoS One 8: e54494.
- Makwana R, Molleman A, Parsons ME (2010) Pharmacological characterization of cannabinoid receptor activity in the rat-isolated ileum myenteric plexus-longitudinal muscle preparation. Br J Pharmacol 159(8): 1608-1622.
- Ijiri Y, Kato R, Sadamatsu M, Takano M, Okada Y, et al. (2014) The effect of Capsaicin on circulating biomarkers, soluble tumor necrosis factor and soluble tumor necrosis factor-receptor-1 and -2 levels in vivo using lipopolysaccharide-treated mice. Toxicology Reports 1: 1062-1067.
- Kriegler M, Perez C, DeFay K, Albert I, Lu SD (1988) A novel form of TNF/ cachectin is a cell surface cytotoxic transmembrane protein: ramifications for the complex physiology of TNF. Cell 53(1): 45-53.
- Idriss HT, Naismith JH (2000) TNF alpha and the TNF receptor superfamily: structure-function relationship(s). Microsc Res Tech 50(3): 184-195.
- Lien E, Liabakk NB, Johnsen AC, Nonstad U, Sundan A, et al. (1995) Polymorphonuclear granulocytes enhance lipopolysaccharide-induced soluble p75 tumor necrosis factor receptor release from mononuclear cells. Eur J Immunol 25(9): 2714-2717.
- Ijiri Y, Kato R, Sadamatsu M, Takano M, Okada Y, et al. (2014) Chronological changes in circulating levels of soluble tumor necrosis factor receptors 1 and 2 in rats with carbon tetrachloride-induced liver injury. Toxicology 316:55-60.





