AT2 receptor: Its role in obesity associated hypertension
Quaisar Ali* and Tahir Hussain*
*Heart and Kidney Institute, Department of Pharmacological and Pharmaceutical Sciences,University of Houston, Houston, TX 77204.
#Department of Pharmacal Sciences, Harrison School of Pharmacy, Auburn University, Auburn AL 36849.
*Corresponding Author
Tahir Hussain, PhD,
Head and Professor, Department of Pharmacal Sciences,
4306F Walker Building, Auburn University, Auburn, AL 36849,
E-mail: tzh0015@auburn.edu
Article Type: Research Article
Received: July 23, 2012;Accepted: September 11, 2012;Published: October 16, 2012
Citation: Tahir Hussain (2012) AT2 receptor: Its role in obesity associated hypertension. Int J Clin Pharmacol Toxicol. 1(1), 15-19, doi: dx.doi.org/10.19070/2167-910X-120003
Copyright: Tahir Hussain© 2012. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
The renin-angiotensin system (RAS) is a hormonal cascade that acts together to regulate blood pressure. Angiotensin II (Ang II) is the major octapeptide of RAS and mediates its cellular and physiological actions by acting on AT1 and AT2 receptor. Most of the cellular and physiological actions of Ang II such as cellular growth and proliferation, vasoconstriction, antinatriuresis and increase in blood pressure are mediated via AT1 receptor. The functions associated with the AT2 receptors are less studied, in part, due to its lower expression in adult tissues. However, AT2 receptor has been suggested as functional antagonist of AT1 receptors and thereby opposes the actions of Ang II mediated via AT1 receptor. Thus, the activation of AT2 receptors has been shown to cause vasodilatation, natriuresis and decrease in blood pressure. After the discovery of the AT2 receptor in various parts of the kidney, including in proximal tubules, there has been an interest in establishing a link between the renal AT2 receptor, renal Na-excretion and blood pressure regulation. Earlier, we have reported that activation of renal AT2 receptors increases urinary Na excretion in obese Zucker rats, in part via inhibiting Na+/K+- ATPase (NKA) activity and stimulating nitric oxide/cGMP pathway in the proximal tubules. An impaired pressure natriuresis and increased AT1 receptor function is believed to be the cause of hypertension in obese Zucker rats and other animal models of obesity. In this review, we are focussing on the role of renin angiotensin system especially AT2 receptors in obesity associated hypertension.
2.Prevalence
3.Pathological triad of obesity, diabetes and hypertension
4.AMechanism of obesity associated hypertension
5.Obesity associated hypertension
5.1 Role of sympathetic nervous system
5.2 Role of atrial natriuretic peptide
5.3 Role of renin angiotensin system
5.4 Overview of renin angiotensin system
6.Components of kidney renin angiotensin system
6.1 Angiotensinogen (AGT)
6.2 Renin and (pro) renin receptor [P] RR
6.3 Angiotensin II
6.4 ACE/ACE2
6.5 Mas receptor
6.6 Angiotensin III
6.7 Angiotensin IV/AT4 receptor
6.8 AT1 receptor
7.AT2 receptor
7.1 General characteristics
7.2 Signal transduction
7.3 Expression
8.Physiological Function
8.1 AT2 receptor opposes AT1 receptor function
8.2 AT2 receptor and Na-excretion
8.3 AT2 receptor and blood pressure
9.Current therapeutic target
10.Acknowledgment
11.References
Introduction
Obesity is defined as having a very high amount of body fat in relation to lean body mass, or body mass index (BMI). Person having BMI of >30 is considered obese. It is one of the most important nutritional disorders worldwide. It has become a global epidemic and is particularly true for the United States, where approximately 300 000 deaths each year are associated with being overweight and obese.
Prevalence
There is a dramatic increase in the prevalence of obesity in the United States within the last decade. In 1996 no states had obesity prevalence rates above 18%, whereas in 2008, almost all the states had obesity prevalence rates of 29% and 23 states are already in obese category (Source: Center for Disease Control and Prevention). Obesity is associated with an increased risk of hypertension and diabetes.
Pathological triad of obesity, diabetes and hypertension
Obesity leads to endothelial dysfunction and impairment of renal function that contribute to the maintenance and development of hypertension [20]. Obesity also increases fat mass in the body. These fat cells release a novel protein called PEDF (pigment epithelium-derived factor). PEDF is released into the bloodstream and causes the muscle and liver to become desensitized to insulin [22]. This results in increase in glucose because the pancreas then produces more insulin to counteract these negative effects. This is one of the mechanism by which obesity leads to diabetes. Moreover, adipose tissue also secretes a large number of cytokines in addition to leptin that modulate glucose metabolism and insulin action. These cytokines also induce suppressor of cytokine signaling-3 (SOCS-3), an intracellular signaling molecule that impairs the signaling of both leptin and insulin and are elevated in obesity [21]. This increased glucose in obesity doubles the risk of mortality in hypertensive patients by affecting renal cellular functions including increase in sodium-glucose transport, cellular hypertrophy [78], synthesis of transforming growth factor-β [63] and matrix accumulation [78]. High glucose also directly affects the cardiovascular functions, vasculature, and neuron damage and also increases sympathetic activation, increased Na absorption which eventually causes hypertension. This pathological triad of obesity, diabetes and hypertension is becoming an economic burden on USA. According to the Center for Disease Control and Prevention, it’s costing more than $187 billion dollar each year to treat obesity associated hypertension. Since renin angiotensin system regulates blood pressure so it is the major therapeutic target to treat hypertension. Although, current available drugs like ACE inhibitors and AT1 blockers improve renal/cardiovascular functions in hypertensive patients but they remain ineffective in treating obesity/diabetes related hypertension where desirable blood pressure levels is below 120 mmHg. It is very difficult to achieve lower pressure in obese/diabetic patients and requires combination of three or more drugs.
Mechanism of obesity associated hypertension
There has been a strong positive correlation between weight gain and blood pressure. These obesity related hypertension affects several organs in the body such as heart, vasculature and kidney. The kidney is one such important organ whose function is severely affected by hypertension [30]. The mechanism by which obesity causes hypertension can be attributed to the enhanced sympathetic and renin angiotensin system activity, alteration of intrarenal physical forces, and hyperinsulinemia [37,38]. Obesity leads to excessive tubular absorption of Na and alters kidney function. This leads to increased extracellular blood volume and hence a shift in pressure natriuresis which is believed to be an important mechanism by which obese person develops hypertension [36]. It has also been postulated that increased renal interstitial pressure due to accumulation of subcapsular fat might lead to tubular compression which further leads to more sodium absorption in the proximal tubules thus raising the blood pressure.
Obesity associated hypertension
Sympathetic nervous system (SNS) plays an important role in cardio-renal function. Activation of SNS especially renal sympathetic nerve activity has been linked to the pathogenesis of obesity associated hypertension [39]. Activation of SNS is partly mediated by hyperinsulinemia, angiotensin II, melanocortin 4 receptors and adipokines such as leptin, tumor necrosis factor α and interleukin-6. Binding of leptin to its receptors in the brain regions activates neuronal pathways that reduces appetite and increases SNS activity leading to an increase in blood pressure. Mutation in the leptin receptor leads to exaggerated plasma leptin which causes early onset of obesity. Several studies suggest a link between adipose tissue and exaggerated SNS activity in muscles and kidneys of normotensive humans [39]. Pharmacological blockade of α and β adrenergic receptors lowers the blood pressure in obese subjects by at least 50-60%. Moreover, renal denervation cause natriuresis and decrease in blood pressure. These observations suggest that increased SNS activity contributes to the development of hypertension in obesity.Hyperinsulinemia in obesity contribute to overactivation of SNS in different tissues including kidney [62]. However the role of insulin resistance in the development of hypertension is somewhat controversial. There is one study wherein aspirin treatment improved insulin resistance but did not prevent the development of hypertension in high-fat fed dogs.
Renal glomeruli contains peptides called atrial natriuretic factor (ANP) that plays an important role in regulating sodium homeostasis [11], fluid balance, vasodilatation and blood pressure. In the kidney, ANP opposes the actions of Ang II via AT1R and causes natriuresis. Low levels of ANP have been observed in obese people and are suggested as one of the mechanisms for obesity-related hypertension [73]. The mechanism associated with the decreased ANP levels in obesity is attributed to upregulated natriuretic peptide clearance receptors (NPR-C) which basically removes natriuretic peptides from the circulation [16]. Reduced ANP function on natriuresis has been reported in obese Zucker rats [5].
Renin angiotensin system (RAS) is a very important hormonal regulator of sodium homeostasis in the kidney [13]. RAS was believed to be an antinatriuretic but recent data suggest that it has both natriuretic and antinatriuretic components. However, during obesity associated hypertension there is an increased RAS activity which shifts the sodium balance from pronatriuretic to an antinatriuretic direction resulting in increased sodium absorption leading to increase in blood pressure. Moreover increased RAS also elevates plasma aldosterone [60] which again leads to an abnormal Na-reabsorption and elevation of arterial pressure. Increased RAS activity has been implicated in the etiology of obesity associated hypertension because blockade of RAS has been implicated as a therapeutic strategy in the management of obesity associated hypertension. Here increased RAS activity is mostly taken in terms of increased renin and/or AT1R function.
According to Guyton’s theory, RAS was considered as circulating endocrine system that regulates blood pressure and Na-homeostasis. The discovery of RAS components in different tissues including brain, heart, vasculature, adipose tissue, gonads, pancreas, placenta, and kidney demonstrates the local/tissue production of Ang II [58]. The tissue RAS plays an important role in normal physiological processes and has been implicated in pathophysiological conditions such as hypertension, congestive heart failure and cardiovascular hypertrophy [19, 47]. The present view of RAS is very complex and is a group of related hormones that act together to regulate blood pressure [68]. When the blood pressure drops for any reasons, special cells in the kidney called juxta-glomerular cells detect those changes and release renin into the blood stream. Renin floats around and converts inactive forms of angiotensinogen into angiotensin I. Angiotensin converting enzyme (ACE) converts inactive angiotensin I into angiotensin II. Ang I and Ang II are further converted into Ang 1-9 and Ang 1-7 by ACE2. Ang 1-9 gets converted to Ang 1-7 by ACE and acts on Mas receptors. Angiotensin II is the most important peptide of RAS and produces its effect by binding onto AT1 and AT2 receptors [15].Ang II via AT1 receptors causes vasoconstriction, salt and water retention, promotes cell growth, releases aldosterone, activates SNS and all these altogether leads to an increase in blood pressure. The effects of AT2 receptor activation are the opposite of those mediated through AT1 receptors [10,25]. Ang II via AT2 receptor promotes vasodilatation and inhibits cellular growth (figure 1). Numerous studies indicate that AT2 receptor has a potential role in blood pressure and natriuresis [32,34,69].
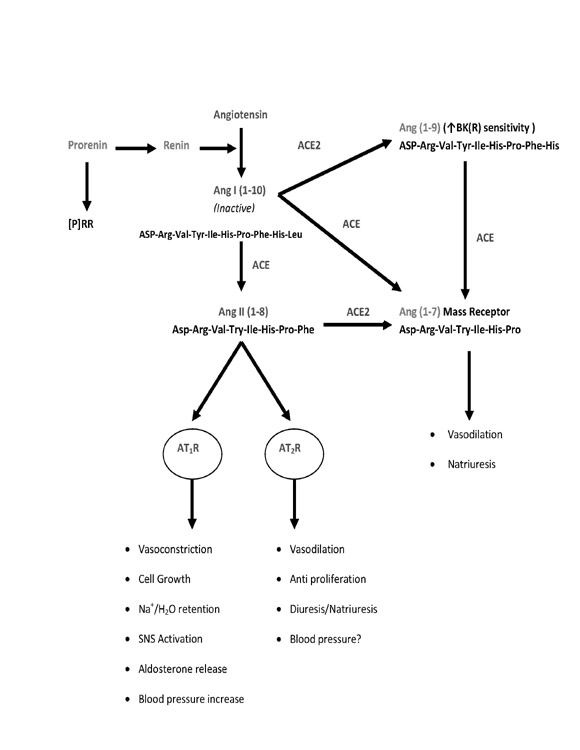

Figure 1: Schematic presentation of renin angiotensin system.
Components of kidney renin angiotensin system
AGT is a glycoprotein consisting of 452 aminoacids. AGT is synthesized in several tissues including liver, heart, blood vessels, adipose tissues and kidney. Renin converts this inactive angiotensinogen into angiotensin I. Increased expression of angiotensinogen gene is observed in plasma samples of hypertensive rats. The increased activity of this gene might lead to more Ang II formation and may cause more renal and cardiovascular damage [61].
Renin is a key enzyme of RAS and is produced from the juxtaglomerular apparatus of the kidney. Renin is considered as a rate limiting enzyme in Ang II production as it converts inactive angiotensinogen into angiotensin I. In addition to enzymatic action, renin and pro-renin also acts as ligands for two receptors leading to cellular responses. The first is the mannose-6-phosphate (M6P) receptor which binds and internalizes both renin and prorenin and hence is called a clearance receptor. The second receptor is the specific (pro) renin receptor ([P]RR), the activation of which initiates downstream signaling cascades [55]. [P]RR is made up of 350 amino acids and consists of single transmembrane domain. Since prorenin is an inactive form of renin, it undergoes proteolytic and non proteolytic activation which leads to the increased activity of the receptor. The binding of this active form of renin to [P]RR decreases its activation energy and leads to the phosphorylation of mitogen-activated protein kinases (MAP kinase p44/42 and extracellular regulated kinases 1/2 (ERK1/2). The phosphorylation of these kinases leads to an increase in plasminogen activator inhibitor 1 and enhanced expression of transforming growth factor (TGF) β1. This result in synthesis of fibronectin and collagen 1 which is important in regulating actin filament dynamics, maintenance of cell structure, growth, movement and cell death [54]. Several studies suggest that overexpression of [P]RR leads to increased blood pressure and aldosterone secretion. As a result of which prorenin receptor inhibitors like allikskerin are used as a therapeutic target to treat high blood pressure [54].
Angiotensin II (Asp-Arg-Val-Tyr-Ile-His-Pro-Phe) is the most important hormone of RAS and produces its effect by binding onto AT1 and AT2 receptor [75]. Most of the actions of Ang II are mediated via AT1 receptors because these are abundant as compared to AT2 receptor. Ang II via AT1 receptor increases peripheral vascular resistance and increased blood pressure.
ACE is an important enzyme of RAS as it converts Ang I to Ang II. ACE also converts Ang 1-7 into smaller angiotensin fragments thereby reversing the vasodilatory effect of Ang 1-7. ACE is considered a pro-hypertensive enzyme because it generates Ang II and also inhibits the peptides such as bradykinin, responsible for vasodilatation [24,61]. ACE2 is a recently discovered enzyme which has 42% structural resemblance to ACE but implicated in reducing the actions of ACE. ACE2 converts Ang II to Ang 1-7 which acts on Mas receptor and causes vasodilation and natriuresis [17]. This enzyme also converts Ang I to Ang 1-9, however the affinity for ACE2 for Ang II is 400-fold higher than Ang I. Since ACE and ACE2 leads to the generation of peptides which has nearly opposite function, a novel concept has been proposed wherein imbalance between ACE/ACE2 could result in different functions. For example increased ACE activity concomitant with reduced ACE2 activity would lead to generation of peptides which would cause more vasoconstriction and vice versa. This balance of ACE/ACE2 in the regulation of different components of RAS is novel target to treat hypertension and renal damage [46].
Ang 1-7 (Asp-Arg-Val-Tyr-Ile-His-Pro) is formed from Ang II by the action of ACE2 and is the agonist for Mas receptor. The association of Ang 1-7, ACE2 and Mas receptor forms a separate branch of renin-angiotensin system called the ACE2/Ang 1-7/Mas axis [23]. The physiological effects mediated by Ang 1-7 is opposite to that of Ang II acting via AT1 receptor.
Ang 1-7 by acting on Mas receptor causes vasodilatation and antiproliferation. Several studies suggest that ACE2/Ang 1-7/Mas axis interacts with the AT1 and AT2 receptor stimulation. For example, it has been shown that stimulation of Mas receptor inhibits the AT1 mediated regulation of ERK1/2 activity which was reversed by Mas receptor antagonist [46]. However, the interaction between AT2 receptor mediated signaling cascade and the ACE2/Ang 1-7/Mas axis is still not known.
Ang III (Arg-Val-Tyr-Ile-His-Pro-Phe) is formed from Ang II by the action of aminopeptidase A. The physiological effects of Ang III are similar to that of Ang II but are less potent. Infusion of Ang III is known to increase BP and intracerebroventricular injection of Ang III is known to increase thirst, vasopressin release and hypertension in animal models [61]. Recent studies suggest that Ang III might be the preferred agonist for AT2 receptor.Infusion of Ang III produces natriuresis via AT2 receptor in AT1 blocked rats [57].
The receptor for Ang IV (Val-Tyr-Ile-His-Pro-Phe) is known as insulin-regulated aminopeptidase (IRAP) or AT4 receptor. IRAP/AT4 is a zinc-bound metalloenzyme attached to the transmembrane domain and their translocation to the cytosol is regulated by insulin. It has a molecular mass of 165 kDa and is made up of 1025 amino acid [2]. Since IRAP is an endopeptidase, it cleaves substrates at the N-terminal of cysteine and leucine amino acids. Ang IV produces its effect by inhibiting activity of IRAP/AT4. This might be one of the mechanism by which Ang IV binds to AT4/IRAP and reduce the cleavage of important peptides and prolong their actions [61]. IRAP/AT4 receptor has a role in maintaining homeostasis during pregnancy by cleaving and inactivating Ang III, oxytocin and vasopressin. The expression of IRAP/AT4 receptor is seen in heart, muscles, liver, spleen, colon and kidney. In kidney these receptors are restricted to proximal tubules, glomerulus, thick ascending loop and collecting ducts [41]. The physiological function of IRAP/AT4 receptor is believed to be similar to that of AT2 receptor in the sense that they can antagonize the function of AT1 receptor by regulating blood flow and promoting Na-excretion [15,40].
AT1 receptor belongs to the family of G-protein coupled receptors. Human AT1 receptor is made up of 359 amino acids and has almost 95% homology with bovine and rodent AT1receptors [14]. It has an extracellular N-terminus followed by a seven transmembrane domain which is connected by three extracellular and intracellular loops linked to the C-terminus. Ang II binds to the extracellular loop and to the transmembrane domain. Receptor internalization, desensitization and phosphorylation of AT1 receptors are linked to the C-terminus of AT1.
AT2 receptor Current therapeutic target Acknowledgement
AT2 receptor belongs to the family of G-protein cou -pled receptor with a molecular mass of 41,000 kDa [15, 69]. The gene which codes for AT2 receptor is present on the X chromosome and has 34% resem -blance with the protein sequence of AT1 receptor [48,51]. There are five potential N-glycosylation sites on the extracellular surface of AT2 receptor [15].
Although, AT2 receptor belongs to the family of G-protein coupled receptor (GPCR), but the entire signaling cascade through G-protein coupling is not known [43]. It is suggested that the third loop of this 7-TM receptor is involved in the downstream signal -ing cascade. Some evidence of AT2 signaling comes from studies in COS-7 cell line and neuronal cell line where it has been shown that agonist occupied AT2 receptor stimulates Giα [42,76]. This receptor is dif -ferent as compared to other GPCR as these agonist occupied receptor does not undergo desensitiza -tion. The reason is still not known but it is speculated that the third intracellular loop is short and does not provide enough binding sites for phosphorylation. Stimulation of AT2 receptor leads to an increase in phosphotyrosine phosphatase activity and inhibition of MAP kinase (p42/p44) or ERK1/2 [9,74]. AT2 re-ceptor stimulation also leads to increase in bradykin -in production which via NO/cGMP pathway causes vasodilation [67]. In our laboratory, we have demon -strated that in proximal tubules of obese Zucker rats, acute activation of AT2 receptor inhibits NKA via NO/cGMP pathway and promotes Na excretion [33].
The expression of the AT2 receptor is observed in sev -eral organs like heart, brain, vasculature, testes and kidney. Within the kidneys, proximal tubules, distal tubules, afferent and efferent arterioles express AT2 re -ceptor [12,59]. The AT2 receptor is widely expressed during embryonic stage and gradually decreases after birth [15,26]. AT2 receptor has drawn limited atten -tion mainly due to its low expression [77]. However, AT2 receptors are overexpressed in various patho-physiological and experimental conditions like obe -sity/diabetes, nephrectomy, atherosclerosis, cardiac overload and myocardial infarction. We believe thatthese overexpressed receptors in these pathophysiological conditions might have a protective role in disease conditions.
AT2 receptors have been shown to produce cellular and physiological responses that are opposite to that produced by AT1 receptor. For example AT2 receptor mediates cellular differentiation and apoptosis in various cells/tissues like vascular smooth muscle cells, endothelilal cells whereas AT1 receptor causes cellular hypertrophy and growth [50]. While Ang II via AT2 receptor causes vasodilation via NO/cGMP pathway whereas AT1 receptor causes vasoconstriction [9]. The mechanism by which AT2 receptor opposes the action of AT1 receptor is not clear. However, there are some studies which suggest that AT2 receptor binds directly to the AT1 receptor and antagonizes its function and this antagonism was linked to the heterodimeriza -tion of these receptors in transfected foetal fibroblast and myometrium of pregnanat women [1]. Ang II via activation of AT1 receptor is known to stimulate NKA causing anti-natriuresis while activation of AT2 receptors inhibits NKA and causes sodium excre -tion [32, 34,56]. Further, AT1 receptor stimulatesNKA activity by reducing cellular cAMP contents [45] whereas AT2 receptor increases cGMP generation which via a cGMP-dependent pathway inhibits NKA activity in the proximal tubules of obese rats [33]. Since cGMP is a known inhibitor of phosphodiesterase-3 (PDE-3), an enzyme that degrades cAMP, AT2 receptor, by increasing cGMP could be inhibiting PDE-3, preventing cAMP reduction and there -by reversing AT1-mediated NKA stimulation [18].
Studies on the role of AT2 receptors on renal sodium transport are limited. In-vitro studies suggest that AT2 receptor mediates inhibition of sodium transport in the proximal tubules of rabbit [31]. AT2 receptor knock-out mice show antinatriuretic hypersensitivity to Ang II and a shift in pressure natriuresis curve. Pres -sure natriuresis is the mechanism by which renal func -tion is linked to long-term blood pressure regulation. However it is difficult to predict whether the effects on sodium excretion are due to absence of AT2 receptor activation or due to enhanced AT1 receptor activity [70]. AT2 receptors are over expressed in the proximal tubules of obese Zucker rats. Activation of AT2 receptors inhibit NKA activity in the proximal tubules and promote natriuresis [3, 33]. Infusion of this AT2 receptor agonist does not affect the glomerular filtration rate (GFR) or mean arterial pressure, suggesting that the changes in natriuresis may be linked to the chang -es in tubular sodium transport [33]. It is known that acute activation of renal AT2 receptors promote natriu -resis/diuresis but whether the long-term AT2 receptor activation modulates the tubular sodium transport, leading to a decrease in sodium balance is not known.
The long-term regulation of blood pressure is linked to the ability of kidneys to excrete sufficient sodi -um to maintain normal sodium balance and blood volume [53]. The AT2 receptor is involved in the production of cGMP, NO and prostaglandin F2α thereby playing an important role in renal function, vasodilatation and blood pressure regulation [52,65]. Data from our laboratory and elsewhere sug -gest that AT2 receptor plays a protective role against increase in blood pressure by promoting sodium excretion. The argument that increase in sodiumexcretion due to AT2 receptor activation may shift the blood pressure can be supported by selective inhibi -tion/disrupting AT2 receptor gene in the kidney. AT2 receptor disrupted mice have increased blood pressure compared to the wild type control and there is sus -tained hypersensitivity of blood pressure and sodium excretion to Ang II [69]. In conscious rats, direct stim -ulation of AT2 receptor with its agonist CGP42112A in the presence of AT1 blocker lowers the arterial pres -sure. The studies so far have been focused on the acute stimulation of AT2 receptor by CGP42112A in spon -taneously hypertensive rat (SHR), normotensive or Sprague Dawley (SD) rats [74]. There is no study done to look at chronic activation of AT2 receptor and its role in long-term blood pressure regulation in obese rats.
Obesity associated hypertension and other renal-car -diovascular diseases are the leading causes of death in the United States. It leads to cardiovascular diseases in -cluding renal ischemia and its dysfunction. In obesity associated hypertension the regulatory function of kid -ney is severely disrupted resulting in irregular sodium excretion and retention. While the known therapeutic target such as ACE, renin inhibitors and AT1 receptor blockers have been effective in treating various forms of hypertension, these targets are often not sufficient to achieve blood pressure goals in obesity/diabetes re -lated hypertension. Recently, discovery of AT2 receptors in adult renal tissues has offered the potential for a novel approach in improving renal function and decreasing high blood pressure. An increase in AT2 receptor expression has recently been reported in animal models [34]. The selective activation of AT2 receptor leads to a greater increase in renal sodium excretion in hyperglycemic animals compared to the normalanimals [35]. Since excessive retention of sodium is a factor for developing hypertension in obesity and dia -betes, AT2 receptor may therefore be a potential candidate for treating hypertension in obese individuals. Un -derstanding the role of AT2 receptor in renal function and blood pressure control will provide a new potential target to treat obesity/diabetes related hypertension.
NIHRO1-DK61578
References
- AbdAlla, S., H. Lother, A. M. Abdel-tawab, and U. Quitterer. 2001. The angiotensin II AT2 receptor is an AT1 receptor antago -nist. The Journal of biological chemistry 276:39721-39726.
- Albiston, A. L., S. G. McDowall, D. Matsacos, P. Sim, E. Clune, T. Mustafa, J. Lee, F. A. Mendelsohn, R. J. Simpson, L. M. Connol-ly, and S. Y. Chai. 2001. Evidence that the angiotensin IV (AT(4)) receptor is the enzyme insulin-regulated aminopeptidase. The Journal of biological chemistry 276:48623-48626.
- Ali, Q., and T. Hussain. 2012. AT(2) receptor non-peptide ago-nist C21 promotes natriuresis in obese Zucker rats. Hyperten-sion research : official journal of the Japanese Society of Hyper-tension.
- Aperia, A., U. Holtback, M. L. Syren, L. B. Svensson, J. Fry-ckstedt, and P. Greengard. 1994. Activation/deactivation of re-nal Na+,K(+)-ATPase: a final common pathway for regulation of natriuresis. Faseb J 8:436-439.
- Baylis, C., C. Foulks, L. Samsell, and K. Engels. 1991. Short term natriuretic responses in the conscious Zucker obese rat. Clinical and experimental hypertension. Part A, Theory and practice 13:1153-1167.
- Bharatula, M., T. Hussain, and M. F. Lokhandwala. 1998. An-giotensin II AT1 receptor/signaling mechanisms in the biphasic effect of the peptide on proximal tubular Na+,K+-ATPase. Clin Exp Hypertens 20:465-480.
- Brown, G. P., and J. G. Douglas. 1982. Angiotensin II binding sites on isolated rat renal brush border membranes. Endocrinol-ogy 111:1830-1836.
- Burson, J. M., G. Aguilera, K. W. Gross, and C. D. Sigmund. 1994. Differential expression of angiotensin receptor 1A and 1B in mouse. The American journal of physiology 267:E260-267.
- Carey, R. M., Z. Q. Wang, and H. M. Siragy. 2000. Role of the angiotensin type 2 receptor in the regulation of blood pressure and renal function. Hypertension 35:155-163.
- Carey, R. M., Z. Q. Wang, and H. M. Siragy. 2000. Update: role of the angiotensin type-2 (AT(2)) receptor in blood pressure regulation. Current hypertension reports 2:198-201.
- Chevalier, R. L., D. G. Muchant, B. A. Thornhill, D. C. Bel-monte, R. A. Pence, and A. J. Baertschi. 1996. Regulation of the renal response to atrial natriuretic peptide by sodium intake in preweaned rats. Pediatric research 40:294-299.
- Clauser, E., K. M. Curnow, E. Davies, S. Conchon, B. Teutsch, B. Vianello, C. Monnot, and P. Corvol. 1996. Angiotensin II re-ceptors: protein and gene structures, expression and potential pathological involvements. European journal of endocrinology / European Federation of Endocrine Societies 134:403-411.
- Crowley, S. D., and T. M. Coffman. 2007. In hypertension, the kidney rules. Current hypertension reports 9:148-153.
- Curnow, K. M., L. Pascoe, and P. C. White. 1992. Genetic analysis of the human type-1 angiotensin II receptor. Mol Endo-crinol 6:1113-1118.
- de Gasparo, M., K. J. Catt, T. Inagami, J. W. Wright, and T. Unger. 2000. International union of pharmacology. XXIII. The angiotensin II receptors. Pharmacological reviews 52:415-472.
- Dessi-Fulgheri, P., R. Sarzani, and A. Rappelli. 1998. The natriuretic peptide system in obesity-related hypertension: new pathophysiological aspects. Journal of nephrology 11:296-299.
- Dilauro, M., and K. D. Burns. 2009. Angiotensin-(1-7) and its effects in the kidney. TheScientificWorldJournal 9:522-535.
- Dousa, T. P. 1999. Cyclic-3’,5’-nucleotide phosphodiesterase isozymes in cell biology and pathophysiology of the kidney. Kid -ney international 55:29-62.
- Dzau, V. 2005. The cardiovascular continuum and renin-angiotensin-aldosterone system blockade. Journal of hyperten-sion. Supplement : official journal of the International Society of Hypertension 23:S9-17.
- Elmarakby, A. A., and J. D. Imig. 2010. Obesity is the major contributor to vascular dysfunction and inflammation in high-fat diet hypertensive rats. Clin Sci (Lond) 118:291-301.
- Emanuelli, B., P. Peraldi, C. Filloux, C. Chavey, K. Freidinger, D. J. Hilton, G. S. Hotamisligil, and E. Van Obberghen. 2001. SOCS-3 inhibits insulin signaling and is up-regulated in response to tumor necrosis factor-alpha in the adipose tissue of obese mice. The Journal of biological chemistry 276:47944-47949.
- Famulla, S., D. Lamers, S. Hartwig, W. Passlack, A. Horrighs, A. Cramer, S. Lehr, H. Sell, and J. Eckel. 2011. Pigment epitheli-um-derived factor (PEDF) is one of the most abundant proteins secreted by human adipocytes and induces insulin resistance and inflammatory signaling in muscle and fat cells. Int J Obes (Lond) 35:762-772.
- Ferrario, C. M., J. Jessup, P. E. Gallagher, D. B. Averill, K. B. Brosnihan, E. Ann Tallant, R. D. Smith, and M. C. Chappell. 2005. Effects of renin-angiotensin system blockade on renal angioten-sin-(1-7) forming enzymes and receptors. Kidney international 68:2189-2196.
- Fleming, I. 2006. Signaling by the angiotensin-converting enzyme. Circulation research 98:887-896.
- Gallinat, S., S. Busche, M. K. Raizada, and C. Sumners. 2000. The angiotensin II type 2 receptor: an enigma with multiple variations. American journal of physiology. Endocrinology and metabolism 278:E357-374.
- Grady, E. F., L. A. Sechi, C. A. Griffin, M. Schambelan, and J. E. Kalinyak. 1991. Expression of AT2 receptors in the developing rat fetus. The Journal of clinical investigation 88:921-933.
- Greger, R. 2000. Physiology of renal sodium transport. Am J
- Griendling, K. K., and R. W. Alexander. 1993. The angiotensin ( AT1) receptor. Seminars in nephrology 13:558-566.
- Guo, D. F., Y. L. Sun, P. Hamet, and T. Inagami. 2001. The angiotensin II type 1 receptor and receptor-associated proteins. Cell research 11:165-180.
- Guyton, A. C., and T. G. Coleman. 1999. Quantitative analysis of the pathophysiology of hypertension. 1969. Journal of the American Society of Nephrology : JASN 10:2248-2258.
- Haithcock, D., H. Jiao, X. L. Cui, U. Hopfer, and J. G. Douglas. 1999. Renal proximal tubular AT2 receptor: signaling and trans -port. Journal of the American Society of Nephrology : JASN 10 Suppl 11:S69-74.
- Hakam, A. C., and T. Hussain. 2006. Angiotensin II AT2 receptors inhibit proximal tubular Na+-K+-ATPase activity via a NO/cGMP-dependent pathway. American journal of physiology. Re-nal physiology 290:F1430-1436.
- Hakam, A. C., and T. Hussain. 2006. Angiotensin II type 2 receptor agonist directly inhibits proximal tubule sodium pump activity in obese but not in lean Zucker rats. Hypertension 47:1117-1124.
- Hakam, A. C., and T. Hussain. 2005. Renal angiotensin II type-2 receptors are upregulated and mediate the candesartan-induced natriuresis/diuresis in obese Zucker rats. Hypertension 45:270-275.
- Hakam, A. C., A. H. Siddiqui, and T. Hussain. 2006. Renal an-giotensin II AT2 receptors promote natriuresis in streptozotocin-induced diabetic rats. American journal of physiology. Renal physiology 290:F503-508.
- Hall, J. E. 2003. The kidney, hypertension, and obesity. Hy-pertension 41:625-633.
- Hall, J. E., M. W. Brands, and J. R. Henegar. 1999. Mecha-nisms of hypertension and kidney disease in obesity. Annals of the New York Academy of Sciences 892:91-107.
- Hall, J. E., E. D. Crook, D. W. Jones, M. R. Wofford, and P. M. Dubbert. 2002. Mechanisms of obesity-associated cardiovascu-lar and renal disease. Am J Med Sci 324:127-137.
- Hall, J. E., A. A. da Silva, J. M. do Carmo, J. Dubinion, S. Hamza, S. Munusamy, G. Smith, and D. E. Stec. 2010. Obesity-induced hypertension: role of sympathetic nervous system, leptin, and melanocortins. The Journal of biological chemistry 285:17271-17276.
- Hamilton, T. A., R. K. Handa, J. W. Harding, and J. W. Wright. 2001. A role for the angiotensin IV/AT4 system in mediating natriuresis in the rat. Peptides 22:935-944.
- Handa, R. K., L. T. Krebs, J. W. Harding, and S. E. Handa. 1998. Angiotensin IV AT4-receptor system in the rat kidney. The Amer-ican journal of physiology 274:F290-299.
- Hayashida, W., M. Horiuchi, and V. J. Dzau. 1996. Intracellular third loop domain of angiotensin II type-2 receptor. Role in mediating signal transduction and cellular function. The Journal of biological chemistry 271:21985-21992.
- Horiuchi, M., M. Akishita, and V. J. Dzau. 1999. Recent pro-gress in angiotensin II type 2 receptor research in the cardiovas-cular system. Hypertension 33:613-621.
- Houillier, P., R. Chambrey, J. M. Achard, M. Froissart, J. Pog-gioli, and M. Paillard. 1996. Signaling pathways in the biphasic effect of angiotensin II on apical Na/H antiport activity in proxi-mal tubule. Kidney international 50:1496-1505.
- Inagami, T., Y. Kambayashi, T. Ichiki, S. Tsuzuki, S. Eguchi, and T. Yamakawa. 1999. Angiotensin receptors: molecular biology and signalling. Clinical and experimental pharmacology & physi-ology 26:544-549.
- Iwai, M., and M. Horiuchi. 2010. Role of the ACE2/angio-tensin1-7/Mas axis in the cardiovascular system. Hypertension research : official journal of the Japanese Society of Hyperten-sion 33:1108-1109.
- Johnston, C. I. 1994. Tissue angiotensin converting enzyme in cardiac and vascular hypertrophy, repair, and remodeling. Hy -pertension 23:258-268.
- Kambayashi, Y., S. Bardhan, K. Takahashi, S. Tsuzuki, H. Inui, T. Hamakubo, and T. Inagami. 1993. Molecular cloning of a novel angiotensin II receptor isoform involved in phosphotyros-ine phosphatase inhibition. The Journal of biological chemistry 268:24543-24546.
- Matsusaka, T., and I. Ichikawa. 1997. Biological functions of angiotensin and its receptors. Annual review of physiology 59:395-412.
- Millatt, L. J., E. M. Abdel-Rahman, and H. M. Siragy. 1999. Angiotensin II and nitric oxide: a question of balance. Regula-tory peptides 81:1-10.
- Mukoyama, M., M. Nakajima, M. Horiuchi, H. Sasamura, R.E. Pratt, and V. J. Dzau. 1993. Expression cloning of type 2 an-giotensin II receptor reveals a unique class of seven-transmem-brane receptors. The Journal of biological chemistry 268:24539-24542.
- Murphy, T. J., R. W. Alexander, K. K. Griendling, M. S. Runge, and K. E. Bernstein. 1991. Isolation of a cDNA encoding the vas-cular type-1 angiotensin II receptor. Nature 351:233-236.
- Navar, L. G. 1997. The kidney in blood pressure regulation and development of hypertension. The Medical clinics of North America 81:1165-1198.
- Nguyen, G. 2007. The (pro)renin receptor: a new kid in town. Seminars in nephrology 27:519-523.
- Nguyen, G. 2007. The (pro)renin receptor: pathophysiologi-cal roles in cardiovascular and renal pathology. Current opinion in nephrology and hypertension 16:129-133.
- Padia, S. H., N. L. Howell, H. M. Siragy, and R. M. Carey. 2006. Renal angiotensin type 2 receptors mediate natriuresis via an- giotensin III in the angiotensin II type 1 receptor-blocked rat. Hypertension 47:537-544.
- Padia, S. H., B. A. Kemp, N. L. Howell, M. C. Fournie-Zaluski, B. P. Roques, and R. M. Carey. 2008. Conversion of renal angio-tensin II to angiotensin III is critical for AT2 receptor-mediated natriuresis in rats. Hypertension 51:460-465.
- Paul, M., A. Poyan Mehr, and R. Kreutz. 2006. Physiology of local renin-angiotensin systems. Physiological reviews 86:747-803.
- Pieruzzi, F., Z. A. Abassi, and H. R. Keiser. 1995. Expression of renin-angiotensin system components in the heart, kidneys, and lungs of rats with experimental heart failure. Circulation 92:3105-3112.
- Queisser, N., P. I. Oteiza, H. Stopper, R. G. Oli, and N. Schupp. 2011. Aldosterone induces oxidative stress, oxidative DNA dam-age and NF-kappaB-activation in kidney tubule cells. Molecular carcinogenesis 50:123-135.
- Ribeiro-Oliveira, A., Jr., A. I. Nogueira, R. M. Pereira, W. W. Boas, R. A. Dos Santos, and A. C. Simoes e Silva. 2008. The renin-angiotensin system and diabetes: an update. Vascular health and risk management 4:787-803.
- Rocchini, A. P. 1995. Insulin resistance, obesity and hyper-tension. The Journal of nutrition 125:1718S-1724S.
- Rocco, M. V., Y. Chen, S. Goldfarb, and F. N. Ziyadeh. 1992. Elevated glucose stimulates TGF-beta gene expression and bio-activity in proximal tubule. Kidney international 41:107-114.
- Ruiz, O. S., Y. Y. Qiu, L. J. Wang, and J. A. Arruda. 1995. Regu-lation of the renal Na-HCO3 cotransporter: IV. Mechanisms of the stimulatory effect of angiotensin II. Journal of the American Society of Nephrology : JASN 6:1202-1208.
- Sasaki, K., Y. Yamano, S. Bardhan, N. Iwai, J. J. Murray, M. Hasegawa, Y. Matsuda, and T. Inagami. 1991. Cloning and ex-pression of a complementary DNA encoding a bovine adrenal angiotensin II type-1 receptor. Nature 351:230-233.
- Schelling, J. R., and S. L. Linas. 1994. Angiotensin II-de-pendent proximal tubule sodium transport requires receptor-mediated endocytosis. The American journal of physiology 266:C669-675.
- Siragy, H. M. 2000. AT(1) and AT(2) receptors in the kidney: role in disease and treatment. American journal of kidney dis-eases : the official journal of the National Kidney Foundation 36:S4-9.
- Siragy, H. M. 2000. The role of the AT2 receptor in hyperten -sion. American journal of hypertension 13:62S-67S.
- Siragy, H. M., and R. M. Carey. 1999. Protective role of the angiotensin AT2 receptor in a renal wrap hypertension model.Hypertension 33:1237-1242.
- Siragy, H. M., T. Inagami, T. Ichiki, and R. M. Carey. 1999. Sus-tained hypersensitivity to angiotensin II and its mechanism in mice lacking the subtype-2 (AT2) angiotensin receptor. Proceed -ings of the National Academy of Sciences of the United States of America 96:6506-6510.
- Thekkumkara, T. J., R. Cookson, and S. L. Linas. 1998. An-giotensin (AT1A) receptor-mediated increases in transcellular sodium transport in proximal tubule cells. The American journal of physiology 274:F897-905.
- Thomas, W. G. 1999. Regulation of angiotensin II type 1 (AT1)receptor function. Regulatory peptides 79:9-23.
- Wang, T. J., M. G. Larson, D. Levy, E. J. Benjamin, E. P. Leip, P. W. Wilson, and R. S. Vasan. 2004. Impact of obesity on plasma natriuretic peptide levels. Circulation 109:594-600.
- Widdop, R. E., E. S. Jones, R. E. Hannan, and T. A. Gaspari. 2003. Angiotensin AT2 receptors: cardiovascular hope or hype? British journal of pharmacology 140:809-824.
- Wolf, G., U. Butzmann, and U. O. Wenzel. 2003. The renin-angiotensin system and progression of renal disease: from hemodynamics to cell biology. Nephron. Physiology 93:P3-13.
- Zhang, J., and R. E. Pratt. 1996. The AT2 receptor selectively associates with Gialpha2 and Gialpha3 in the rat fetus. The Jour-nal of biological chemistry 271:15026-15033.
- Zhuo, J., K. Song, P. J. Harris, and F. A. Mendelsohn. 1992. In vitro autoradiography reveals predominantly AT1 angiotensin II receptors in rat kidney. Renal physiology and biochemistry 15:231-239.
- Ziyadeh, F. N., K. Sharma, M. Ericksen, and G. Wolf. 1994. Stimulation of collagen gene expression and protein synthesis in murine mesangial cells by high glucose is mediated by auto -crine activation of transforming growth factor-beta. The Journal of clinical investigation 93:536-542.





