Renaming Bombyx mori Chemosensory Proteins
Jean-François Picimbon
Overseas Talent and Taishan Scholar (NO.tshw20091015), Functional Genomics and Proteomics of Chemical Ecology, Biotechnological Research Center, Shandong Academy of Agricultural Sciences, 202 North Industrial Road, 250100 Jinan, China.
*Corresponding Author
Jean-François Picimbon,
Overseas Talent and Taishan Scholar (NO.tshw20091015),
Functional Genomics and Proteomics of Chemical Ecology,
Biotechnology Research Center, Shandong Academy of Agricultural Sciences,
202 North Industrial Road, 250100 Jinan, China.
Tel: 0086-531-83175350; Fax: 0086-531-83178156
E-mail: jfpicimbon@gmail.com
Article Type: Editorial
Received: May 07, 2014; Published: May 29, 2014
Citation: Jean-François Picimbon (2014) Renaming Bombyx mori Chemosensory Proteins, Int. J. Bioorg. Chem. Mol. Biol. 2(2e), 1-4. doi:dx.doi.org/10.19070/2332-2756-140005e
Copyright: © 2014 Jean-François Picimbon. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
This is a concise critical editorial about the functions described for a group of proteins from the family of Chemosensory Proteins (CSPs), particularly functions related to specific binding of odor chemosensory molecules. CSPs are small soluble proteins of a size comprised between 10-14 kDa and a flexible structure made of four Cysteine residues and six alpha-helices [1]. They are found to bind a large repertoire of ligand molecules mainly in the fatty acid category in most of all insect species. A specific function in binding odor chemosensory ligands has been proposed on the basis of the presence of these proteins in the sensory lymph of locusts. This finding has given the name of the whole protein superfamily, CSPs [2].
As we progress into the full-description of the CSP gene family in an increasing number of insects, in particular in moths, we bring more and more evidence that most of the CSPs play no or little role in olfaction. The findings that the cockroach p10 protein is highly expressed in regenerating tissues such as legs and that the same protein is abundantly expressed in the adult fully grown antennae are two facts that have been eluded perhaps too quickly [3,4]. From beginning, molecular studies analysing tissue-specific gene expression and developmental profiling in moths such as the silkworm (Bombyx mori) and the tobacco budworm (Heliothis virescens) bring one of the more important points to discuss about the function of this protein family: CSP genes are expressed throughout the whole insect body in both internal and external tissues at various developmental stages [5,6]. Applying cDNA microarray, tag EST and transcriptomic technologies for moth CSP research confirmed this breakthrough pioneer idea [7,9]. Studying in details the CSP protein family in other insect species such as locusts and bees led to the same conclusion [10-12]. Finally, even immunocytochemical studies are in support of a non-sensory function for CSPs; immunolocalization of CSPs in both solitary and gregarious locusts shows a pattern much broader than it appears initially. CSPs are found outside the sensory lymph, in the subcuticular space between epidermis and cuticle [13].
CSPs are found everywhere in different forms of the insect and most probably play a very general physiological function and/or retain all types of functions at all stages of the insect development. Many hormones such as juvenile hormone are known to have such a versatile profile [14]. However, to the best of our knowledge, there have been no previous reports of such broadly tuned proteins, except perhaps the odorant-binding protein family.
The first experiment of CSP knock out has been done in bees, revealing that CSP plays a key role in head and thereby tissue formation. However, it could not be established from this experiment whether the CSP has only one function tuned to development and/or retains a dual “chemo-devo” function [15]. In moths, the same CSP could well be involved in tissue development at the pupal stage and in olfactory mechanisms at the adult stage. Campanacci et al. have shown that CSPs from moths are capable of remarkable structural flexibility [16]. Xuan et al. have shown that an enormous repertoire of CSPs exists in the moth pheromone gland through RNA editing processes [17]. However, it has to be taken into consideration that the female pheromone gland of the silkworm moth expresses nearly all BmorCSPs while B. mori produces only two chemosensory ligands, Bombykol and Bombykal [17]. This may be seen as indisputable complementary evidence that CSPs have alternative functions to development and transportation of pheromone and/or odor chemicals. A similar profile of CSPs has been described in the female pheromone gland from the tobacco budworm Heliothis virescens [18]. Curiously enough, in the sweetpotato whitefly Bemisia tabaci, it has been shown that insecticide exposure strongly affects CSP gene expression in particular in females [19]. A companion study analysing twenty CSP genes in the female silkworm moth B. mori demonstrates that only two CSP genes (CSP1 and CSP2) are highly (but not only) expressed in sensory organs and that all other CSP genes essentially express in many non-sensory tissues including epidermis, gut, fat body, thorax and wings [20]. Interestingly, building a fast distance-based phylogenetic tree (RAxML) from a sequence alignment (MUSCLE) comparing Bombyx and Heliothis CSP amino acid full-sequences shows that moth CSPs clearly segregate into tissue-specific orthologous groups with strong bootstrap support (Figure 1). The distribution of tissue-specific orthologous groups is such that non-sensory tissue expression of CSPs came first, before the appearance of a group of CSPs mainly expressed in epidermis, fat body, gut, thorax and wings (EFGTW) and a group of CSPs more abundantly (although not uniquely) expressed in sensory tissues such as the antennae, legs and pheromone gland (ALGPFW). CSPs may have developed in sensory organs simply in line with the appearance and development of new sensory systems over the course of evolution (acquisition of mandibules, expansion of antennae, segmentation of legs, shifts to glandular pheromones of diverse structures, etc). Still, all CSPs retain expression in non-sensory tissues, particularly the gut. BmorCSP3 and HVPG0029 fall out of the tree, suggesting perhaps that the ancestral CSPs were expressed throughout the whole insect body but mainly in the fatty tissue, epidermis and gut. CSPs from all various branches of the tree express in the gut. The gut may be the central organ for CSPs.
In the female silkworm B. mori, hundred to thousand-fold increase in expression of CSP genes is observed in the gut after insecticide treatment, strongly suggesting a role in drug metabolism and tissue detoxification perhaps in relation with cytochrome P450 oxidases [20].
The analysis of tissue-expression profiling of the silkworm B. mori CSPs (BmorCSPs) does not support a function in relation with chemosensing at all (Table 1). In “control” conditions (i.e. in absence of external toxic stimuli), very low expression levels are found for BmorCSP10, BmorCSP13, BmorCSP15 and BmorCSP17. Some CSPs such as BmorCSP4 and BmorCSP19 are mainly found in one single tissue. All other BmorCSPs are variably expressed in all tissues from antennae to gut [20]. BmorCSP4, BmorCSP15 and BmorCSP17 seat together with all other CSP genes, but BmorCSP10, BmorCSP13 and BmorCSP19 have a more distant location from the main CSP group. BmorCSP10 gene is gigantic in size (3 exons-2 introns) and codes for an atypical type of CSP protein (24.65 kDa, 211 amino acids, 22 Proline residues). Applying insecticide increased expression of most of all BmorCSPs including BmorCSP10 in most of all tissues of the insect body. BmorCSP1 and BmorCSP2 are mainly expressed in the antennae but not only. They are also found to be up-regulated in the head and gut. Five BmorCSPs (BmorCSP9, BmorCSP11, BmorCSP12, BmorCSP15 and BmorCSP19) are remarkably upregulated in the gut. BmorCSP13, BmorCSP14 and BmorCSP20 are remarkably up-regulated in the thoracic tissue, legs and fat body, respectively. Some CSPs such as BmorCSP17 are markedly up-regulated only in two different tissues (antennae and wings). However, expression of BmorCSP3, BmorCSP4, BmorCSP7,BmorCSP8 and BmorCSP10 genes is significantly increased in many various tissues [20]. Expression of BmorCSP6 is significantly decreased in the antennae, legs, pheromone gland and epidermis (Table 1) [20]. There is no CSP from the silkworm moth B.mori that is intrinsically highly specific to one single tissue. Most importantly, there is no BmorCSP intrinsically highly specific to the antennae. Most of all tissues from antennae to gut contain nearly all BmorCSPs although at different levels (Figure 1 & Table 1) [20-22].
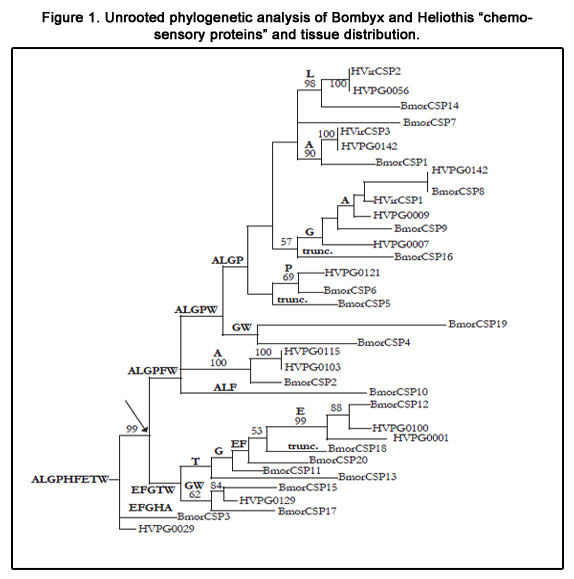
Figure 1. Unrooted phylogenetic analysis of Bombyx and Heliothis “chemosensory proteins” and tissue distribution.
BmorCSPs: BmorCSP1 (AF509239, AJ973410, AK384328, AK383152, AK386791, AK382876, JQ253595-JQ253630, JQ085850- JQ085858, NM_001043587), BmorCSP2 (AF509238, AJ973407, AK386776, AK386789, JQ253631-JQ253667, NM_001043715), BmorCSP3 (AJ973412, AK382932, NM_001043935), BmorCSP4 (JQ253663-JQ253705, NM_001098310), BmorCSP5 (bmb007542), BmorCSP6 (AK385783, NM_001043600), BmorCSP7 (AK382897, NM_001098308), BmorCSP8 (AB243748, AJ973404, AK383175, AK386777, NM_001043598), BmorCSP9 (AB243748, AK383030, AK382874, NM_001043597), BmorCSP10 (AK386835, NM_001043604), BmorCSP11 (AK383425, NM_001043602), BmorCSP12 (AB243754, AK383879, AK378890, NM_001043603), BmorCSP13 (AB243752, AK382964, NM_001043603), BmorCSP14 (AJ973405, AK382964, DQ855516, JQ253712-JQ253781, NM_001043599), BmorCSP15 (NM_001098309), BmorCSP16 (bmb037911), BmorCSP17 (AJ973414, AK383002, NM_001043727), BmorCSP18 (bmb040762), BmorCSP19 (AK382814, NM_001098312), BmorCSP20 (NM_001098311) [20,21]. HvirCSPs: HvirCSP1 (AAM77041, AAV34686), HvirCSP2 (AAM77040), HvirCSP3 (AAM77042), HVPG0001 (ACX53692), HVPG0009 (ACX53700), HVPG0029 (ACX53719), HVPG0037 (ACX53727), HVPG0056 (ACX53745), HVPG0103 (ACX53788), HVPG0115 (ACX53800), HVPG0119 (ACX53804), HVPG0121 (ACX53806), HVPG0129 (ACX53813), HVPG0133 (ACX53817), HVPG0142 (ACX53825). Variant clones such as HVPG0045 (only one Cysteine), HVPG0073 (truncated sequence), HVPG0099 (ten Cysteines) and HVPG0102 (only two Cysteines) were not considered for the analysis (ACX53734, ACX53762, ACX53784, ACX53787). Trunc.: Truncated genes. Numbers above the branches are bootstrap values (>50, 100 bootstrap replicates) [22]. The arrow indicates the position of a switch from non-sensory to sensory expression of moth CSPs. This switch may have happened before the appearance of the major clades of Lepidoptera (more than 150 Mya). Most of all tissue-specific orthologous groups include both Bombyx and Heliothis CSPs. A: Antennae, E: Epidermis, F: Fat body, G: Gut, H: Head, L: Legs, P: Pheromone gland, T: Thorax, W: Wings.
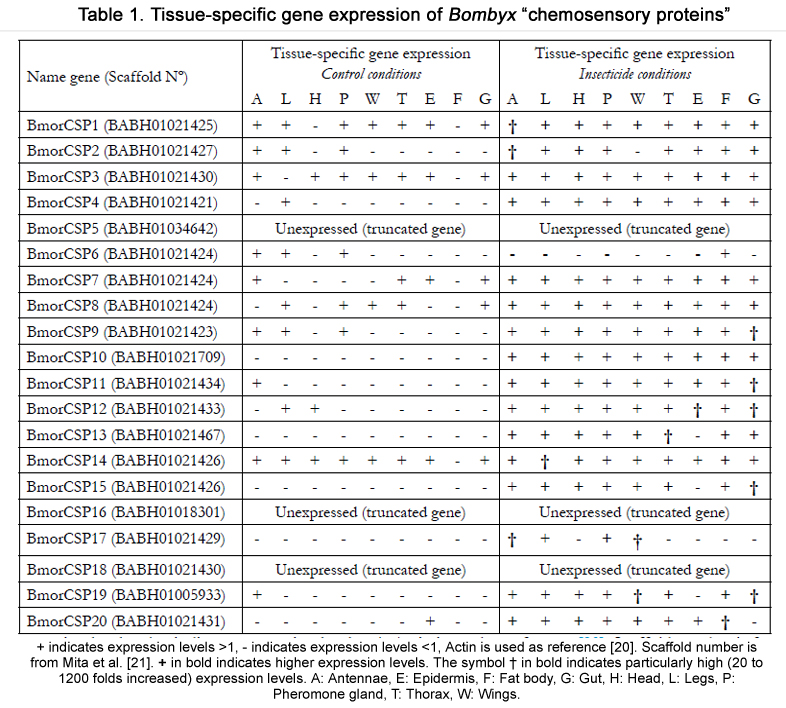

Table 1. Tissue-specific gene expression of Bombyx “chemosensory proteins”.
+ indicates expression levels >1, - indicates expression levels <1, Actin is used as reference [20]. Scaffold number is from Mita et al.[21]. + in bold indicates higher expression levels. The symbol † in bold indicates particularly high (20 to 1200 folds increased) expression levels. A: Antennae, E: Epidermis, F: Fat body, G: Gut, H: Head, L: Legs, P: Pheromone gland, T: Thorax, W: Wings.
Based on these results, it certainly cannot be that CSPs are specifically tuned to chemosensory odor detection. We rather argue that CSPs play a key role in general lipid metabolism. This is a coherent argument to assent to the structure of the binding domain of CSPs (an open-air tube with long aliphatic chains ensheathed), the binding properties of these proteins towards lipids of all kinds, the huge repertoire of BmorCSP variants identified in the pheromone gland, their very general developmental and tissue-expression profiling as well as their insecticide response. It makes consistent the binding of cuticular hydrocarbons to ant CSPs [23]. It also makes consistent the existence of CSPs in aquatic arthropod species such as the water flea Daphnia pulex and the shrimp Artemia franciscana (ABH88167, ABH88166 and ABY62738). Chemosensory molecules detected by fleas and shrimps in the marine environment are necessarily very different than the highly volatile airborne chemosensory desaturated hydrocarbon pheromone odor chemical molecules perceived by insects in the terrestrial environment. In contrast, lipids are common to all arthropods from trilobites to ants and bees.
BmorCSPs, H. virescens, moth, ant, bee, locust, whitefly, flea,shrimp CSPs and most probably also the whole and complete protein family should be renamed accordingly. This may help infer new functions to all CSP proteins, interpret the results in a new way independently of olfactory consideration and inspire science towards the discovery of the true function of this more and more intriguing insect-arthropod protein family.
References
- Picimbon JF (2003). Biochemistry and Evolution of CSP and OBP Proteins. Insect Pheromone Biochemistry and Molecular Biology, the Biosynthesis and Detection of Pheromones and Plant Volatiles, Elsevier Academic Press, San Diego, London, 539-566.
- Angeli S, Ceron F, Scaloni A, Monti M, Monteforti G, et al (1999) Purification, structural characterization, cloning and immunocytochemical localization of chemoreception proteins from Schistocerca gregaria. Eur. J. Biochem. 262:745-754.
- Nomura A, Kawasaki K, Kubo T, Natori S (1992) Purification and localization of p10, a novel protein that increases in nymphal regenerating legs of Periplaneta americana (American cockroach). Int. J. Dev. Biol. 36:391-398.
- Picimbon JF, Leal WS (1999) Olfactory soluble proteins proteins of cockroaches. Insect Biochem. Mol. Biol. 30:973-978.
- Picimbon JF, Dietrich K, Angeli S, Scaloni A, Krieger J, et al. (2000a) Purification and molecular cloning of chemosensory proteins from Bombyx mori. Arch. Insect Biochem. Physiol. 44:120-129.
- Picimbon JF, Dietrich K Krieger J, Breer H (2001) Identity and expression pattern of chemosensory proteins in Heliothis virescens (Lepidoptera, Noctuidae). Insect Biochem. Mol. Biol. 31:1173-1181.
- Gong DP, Zhang H, Zhao P, Lin Y, Xia QY, et al (2007) Identification and expression pattern of the chemosensory protein gene family in the silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 37:266-277.
- Xu YL, He P, Zhang L, Fang SQ, Dong SL, et al . (2009) Large-scale identification of odorant-binding proteins and chemosensory proteins from expressed sequence tags in insects. BMC Genomics 10:632.
- Zhang YN, Jin JY, Jin R, Xia YH, Zhou JJ, et al. (2013) Differential expression patterns in chemosensory and non-chemosensory tissues of putative chemosensory genes identified by transcriptome analysis of insect pest the purple stem borer Sesamia inferens (Walker). PLoS One 8:e69715.
- Picimbon JF, Dietrich K, Breer H, Krieger J (2000b) Chemosensory proteins of Locusta migratoria (Orthoptera : Acrididae). Insect Biochem. Mol. Biol. 30:233-241.
- Forêt S, Wanner KW, Maleszka R (2007) Chemosensory proteins in the honeybee: Insights from the annotated genome, comparative analysis and expression profiling. Insect Biochem. Mol. Biol. 37:19-28.
- Guo W, Wang XH, Ma ZY, Xue L, Han JY, et al. (2011) CSP and Takeout genes modulate the switch between attraction and repulsion during behavioural phase change in the migratory locust. PLoS Genetics 7:e10001291.
- Jin X, Brandazza A, Navarrini A, Ban L, Zhang S, et al. (2005) Expression and immunolocalization of odorant-binding and chemosensory proteins in locusts. Cell. Mol. Life Sci. 62:1156-1166.
- Picimbon JF, Bécard JM, Sreng L, Clément JL, Gadenne C (1995) Juvenile hormone stimulates pheromonotropic brain factor release in the female black cutworm, Agrotis ipsilon. J. Insect Physiol. 41:377-382.
- Maleszka J, Forêt S, Saint R, Maleszka R (2007) RNAi-induced phenotypes suggest a novel role for a chemosensory protein CSP5 in the development of embryonic integument in the honeybee (Apis mellifera). Dev. Genes Evol.217:189-196.
- Campanacci V, Lartigue A, Hällberg BM, Jones TA, Giuici-Orticoni MT, et al. (2003) Moth chemosensory protein exhibits drastic conformational changes and cooperativity on ligand binding. Proc. Natl. Acad. Sci. USA 100:5069-5074.
- Xuan N, Bu X, Liu YY, Yang X, Liu GX, et al. (2014) Molecular evidence of RNA editing in Bombyx chemosensory protein family. PLoS One 9:e86932.
- Vogel H, Heidel AJ, Heckel DG, Groot AT (2010) Transcriptome analysis of the sex pheromone gland of the noctuid moth Heliothis virescens. BMC Genomics 11:29.
- Liu GX, Xuan N, Chu D, Xie HY, Fan ZX, et al. (2014) Biotype expression and insecticide response of Bemisia tabaci chemosensory protein-1. Arch. Insect Biochem. Physiol. 85:137-151.
- Xuan N, Guo X, Xie HY, Lou QN, Lu XB, et al. (2014) Increased expression of CSP and CYP genes in adult silkworm females exposed to avermectins. Insect Sci. doi: 10.1111/1744-7917.12116.
- Mita K, Kasahara M, Sasaki S, Nagayasu Y, Yamada T, et al. (2004) The genome sequence of Silkworm, Bombyx mori. DNA Res. 11:27-35.
- Stamatakis A, Ludwig T, Meier H (2005) RAxML-III: a fast program for maximum likelihood-based interference of large phylogenetic trees. Bioinformatics 2:456-463.
- Ozaki M, Wada-Katsumata A, Fujikawa K, Iwasaki M, Yokohari F, et al. (2005) Ant nestmate and non-nestmate discrimination by a chemosensory sensillum. Science 309:311-314.





