Effect of Influenza-A Virus Infection On Inflammatory gene Expression Profiles Of Leukocyte Concentrate Buffy Coats and Exacerbation of Azthma (Inflammatory Response to Influenzaa Virus Infection)
Al-Ghazal AT1*, Ismail SI2, Al-Umary YI3, Al-Khuzie RF4, Assaf AA5
1 Faculty of Science, Department of Biology, University of Mosul, Iraq.
2 Faculty of Medicine, Department of biochemistry, University of Jordan, Jordan.
3 Faculty of Science, Department of Biology, University of Jordan, Jordan.
4 Faculty of Allied Medical Sciences, Department of Nutrition and food Science, University Applied Sciences and Technology, Iraq.
5 Faculty of Pharmacy, Department of Biopharmaceutics and Clinical Pharmacy, University of Jordan, Jordan.
*Corresponding Author
Abdulrhem T Al-Ghazal,
Faculty of Science, Department of Biology,
University of Mosul, Postal code: 11942, P.O.BOX: 13739, Iraq.
Tel: (00964)07704139591
Fax: 00962 65356746
E-mail: bakurius@yahoo.com
Received: September 14, 2016; Accepted: October 28, 2016; Published: October 31, 2016
Citation: Al-Ghazal AT, Ismail SI, Al-Umary YI, Al-Khuzie RF, Assaf AA (2016) Effect of Influenza-A Virus Infection On Inflammatory gene Expression Profiles Of Leukocyte Concentrate Buffy Coats and Exacerbation of Azthma. (Inflammatory Response to Influenzaa Virus Infection). Int J Virol Stud Res. 4(5), 55-63. doi: http://dx.doi.org/10.19070/2330-0027-160009
Copyright: Al-Ghazal AT© 2016. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
Objectives: Influenza A virus is a major cause of respiratory infections with high rates of morbidity and mortality worldwide. An understanding of how InfluenzaA virus (IAV) modulates host cellular responses is critically important to explore the molecular mechanisms of viral-host interaction. The aims of this study are; to detect changes in the mRNA expression in a panel of inflammatory genes in the leukocyte concentrate Buffy coat of IAV infected patients after 48 hrs of infection. Also to determine the relation between the inflammatory gene expression and asthma in IAV patients.
Methods: Blood from 90 hospital admitted patients suffering from flu within the first 48 hrs of infection was tested for IgM- influenzaA virus. Only eight patients were positive. Leukocytes from the IAV positive patients were tested for a panel of 84 inflammatory genes using real time-PCR array technology.
Results: Only 14 inflammatory genes (IL1B, IL8, IL10, IL13, CCL2, CCL5, CCL7, CXCL1, CXCL10/IP-10, CX3CR1, C5, TNF, ABCF1, and BCL6) shown a significant upregulation fold ranging between between 1.01 and 121.35 fold in all the eight patients with a 100% frequency. The upregulation of IL8, IL 10, IL13, C5, CCL7, CCL5/RANTES, CXCL1 and CCL18mRNA transcription with high significance might suggest that the asthma complication during IAV infection is due to the stimulation of immune response. Six inflammatory genes (CEBPB, CCR1, IL1R1, MIF, CXCL11 and IL9R) shown a decrease in the mRNAexpression with fold ranged between -250.99 and -1.11 compared to control cells after recovery but with variable frequencies indicating a time dependent response.
Conclusion: Our results revealed several alterations of many Leukocyte`s inflammatory gene expressions induced by influenza A virus. Eight of the over expressed genes are involved in asthma complication. All samples are mostly infected with related subtypes of influenza A viruses. These results may help for further analysis of influenza A virus role on host–pathogen interaction.
2.Introduction
3.Materials and Methods
3.1 Patients And Blood Samples
3.2 ELISA Test
3.4 Extraction of mRNA from Patient's Leukocyte
3.5 RNA Quantitation
3.6 RNA Efficiency Test
3.7 Real time- PCR Array
3.8 Real-Time PCR Detection
4.Statistical Analysis
5.Results
5.1 The Over Expression af Inflammatory Genes
5.2 The Down Regulation Of Inflammatory Genes
5.3 Clustering of gene Expression Profile after IAV Infection
6.Discussion
6.1 Clustering of Leukocyte gene Expression Profile after IAV Infection
7.Conclusion
8.Acknowledgements
9.References
Keywords
RT-PCR - Arrays; Influenza A Virus - White Blood Cell Interaction; Inflammatory gene Expression; Cytokines; Cytokines Receptors.
Introduction
Human Influenza A Virus (IAV) causes a highly contagious acute respiratory illness, which can be fatal, particularly in young children and the elderly [1]. For most severe IAV pandemics, the exact contribution of individual viral effect to pathogenicity at molecular levelis still largely unknown. The understanding of such a systematic analysis of IAV infection at the level of transcriptional regulation in the host cells is important for interpretation of these complex genetic changes [2]. IAV is the most able to escape the innate immune response in the airways with the potential consequence of severe respiratory symptoms and systemic inflammation [3]. Viral respiratory infections have a major impact in individuals with chronic airway diseases such as respiratory inflammations, asthma and chronic obstructive pulmonary disease (COPD). In these individuals the clinical response occurs by the effect of the virus on host cell gene expression, the stimulation of antiviral immune response, and the pre-existing of chronic airway inflammation [4]. Viral infections cause changes in gene expression of many pro-inflammatory cytokines and chemokines. Cytokines play a key role in the development and functioning of both the innate and adaptive immune response. They are often secreted by immune cells that have encountered a pathogen such as viruses, thereby activating and recruiting further immune cells to enhance the immune response to the pathogen [5]. Chemokines are essential mediators of inflammation and important for control of viral infections. The profile of chemokine expression was found to have a role in modulating the immune response during viral infection [6]. Previous studies showed that the effect of IAV on the expression of inflammatory genes in response to infection in human cells, contributes to viral pathogenesis, primarily by enabling the virus to disarm the host cell defense system [7, 8]. IAV was found to have the capability of upregulating the expression of a variety of chemokine genes in infected human macrophages or leukocytes leading to the activation of cell signaling pathways and augmentation of inflammation [9, 10]. Tong et al., 2003 showed that the Infection with IAVcan cause changes in the expression of inflammatorygenes in human epithelial cells. Most of these genes were upregulated (2 – 5 fold) including 131 gene (interferon inducible genes, chemokine and cytokine genes, pro-and anti-apoptotic genes, signal transduction and transcription factors, cellular immune response, cell cycle and metabolism genes) and 11 genes were down regulated (Genes of cell membranes, cell adhesion, cell motility, transporter, signaling and transcription) [11].
Real-time-PCR array is an alternative to microarray techniques for the analysis of transcription from multiple genes. The main advantage of real-time PCR array is that it is more sensitive to lowfold changes than other high-throughput assays. In addition, the real-time PCR array technique can detect low quantity of nucleic acid, and has a large dynamic range, and various controls that can be included to ensure accuracy. In contrast to microarray techniques, in real-time PCR array the parameters for each gene can be optimized individually. Moreover, the identity of PCR products can be confirmed through melting curve analysis, restriction endonuclease analysis, or DNA sequencing [12, 13]. The objectives of this study are;to determine the changes in the mRNA expression of a panel of 84 inflammatory genes in the leukocyte concentrate of IAV infected patients after 48 hrs of infection. Also to detect the correlation between the inflammatory gene expression and the exacerbation of asthma after IAV infection.
Eight females hospitalized at Ibn AL-Haytham Hospital in Amman/ Jordan for respiratory tract infections are included to analyze the gene expression changes in leukocytes in response to IAV infection. EDTA-blood (5ml) was collected from patients in the acute phase of the disease (within 48 hrs of infection) with highgrade fever.
Blood samples (2ml) from 90 patients were tested for the viral type using indirectELISA-IgMkit (IBL-Hamburg Corporation) to determine positive IAV patients. The second part of the blood sample (3 ml) is used for RNA extraction. A control blood sample was taken one month after infection from patients showing positive IAV infection to insure a complete recovery and was tested the same way.
The RNA is extracted within 24h ofblood collection as follows: RNA-extraction phase, treat the leukocytes (isolated from the blood Buffy coat) with Trizol reagent (Phenol-guanidin-isothiocyanate); Separation phase, using chloroform; RNA precipitation phase, using Isopropanol Alcohol; washing phase, using 70% ethanol, and dissolving phase; using Rnase-Dnase free water (In vitro gen) [14].
Spectrophotometer (Bio-Rad) analysis is carried out to determine the concentration (μg/ml) of RNA by measuring the optical density (O.D) at λ260 and according to the following equation: RNA concentration = 40 (O.D factor) x 25 (dilution factor) x O.D at λ260 [13]. RNA quality is tested using two methods, spectrophotometer (Bio-Rad) andgel-electrophoresis. In spectrophotometer the O.D260/280 ratio is checked. RNA is considered pure if the O.D ratio is between 1.8 and 2.0.
Before performing the RT PCR-Array, the RNA samples are tested for the efficiency of the reverse transcription to produce the cDNA on the RNA samples (Figure 1) using 1.0μg of random primer, PCR components and master mixture (Promega USA) [15].
Figure 1. Gel Electrophoresis Test for the Efficiency of RNA Samples to Produce the cDNA.
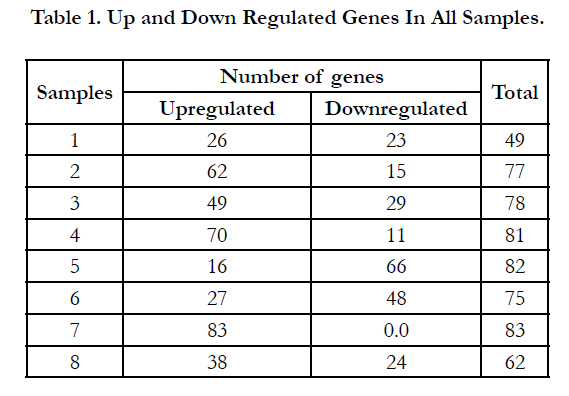

Table 1. Up and Down Regulated Genes In All Samples.
Real time-PCR array were performed to specifically quantitate human cytokine and chemokine mRNA levels in presence or absence of IAV infection to monitor the mRNA expression levels of 84 different cytokines and chemokines in patient`s leukocytes concentrate. Isolated cDNA were synthesized using SABioscience kit. Each cDNA sample was added to RT-qPCR master Mix containing SYBR Green and reference dye (SABioscience). 25μL of cDNA-Master Mix mixture was added to each well across the PCR-arrays (two RT-PCR-array plates for each patient; one during IAV infection and the second for control, after recovery) each PCR-array profiles the expression of 84 pathway-specific geneplus 12 internal controls (SABioscince). Thermocycling conditions were 95°C for 10 min and 40 cycles of (95°C for 15 sec, 60°C for 1 min) using Bio-Rad-iCycler real-time PCR detection systems.
The threshold cycle values (Ct values) are calculated for each well using iQ cycler Bio-Rad software. The data is exported to a blank Excel Spread-sheet for use with SABioscience Data Analysis Template Excel File. Melting curve program (cycle 1, 1 repeat, 95C for 1 min. cycle 2, 1 repeat, 55C for 1 min. cycle 3, 80 repeat, 55C for 10 sec.) is run immediately after the cycling program, and generates a first derivative dissociation curve for each well in the entire plate. The fold-change is calculated for each gene in control and experimental groups as 2(-ΔΔCt). If the fold-change is greater than 1, then the result may be reported as a fold up-regulation. If the fold-change is less than 1, then the negative inverse of the result may be reported as a fold down-regulation.
Statistical Analysis
Since each gene has 16 Ct-values (8 Ct-values with experimental sample and 8 Ct-values with control sample) the difference between the two Ct-values for each gene is analyzed using the Non-parametric Wilcoxon signed ranks-test to calculate the Zvalue and P-value for each gene [16].
Results
Through the period from May 17 to July 8/ 2009, 90 patients were hospitalized at IbnAL Haytham Hospital in Amman/Jordan for respiratory tract infection symptoms. All the 90 patients were subjected to ELISA-IgM test. Only 8 patients gave positive results for IAV.
Influenza A virus infection causes up and down regulation in the expression of a large number of inflammatory genes (Table 2) but only 21 out of the 84 tested cytokines and chemokines genes showed an increase in the mRNA expression level over level of expression on the control cells after recovery. These were classified into four groups based on their frequency (Frq) in the eight different IAV positive patients (genes may be upregulated in more than one or in all 8 samples). As shown in the table 3 the first group includes 14 upregulated genes in all IAV positive patients with 100% frequency which indicates their involvement during cellular response against IAV infection and their ability to stimulate directly or indirectly the immune-inflammatory response.
Eight of the 14 genes (IL8, IL 10, IL13, C5, CCL7, CCL5/ RANTES,CXCL1 and CCL18) are known for their significance in the exacerbation of asthma due to IAV infection [9, 19, 32, 25, 23, 39, 27, 46]. Six of them(IL8,IL10, IL13, CCL7, CCL5/ RANTES, CXCL1) in addition to another four genes of the same group (IL1B, CCL2,TNF, and CX3CR1)were found to play a key role in the cytokine-cytokine receptor interaction pathway [17]. Cytokines IL8 and IL13 were the most highly activated proinflammatory genes products induced in all eight patients with 100% frequency. The virus induced maxima of 121.35 and 72.36 fold increases in the expression of IL8 and IL13 transcription respectively. On the other hand two chemokine genes (CCL4 and CCL18) showed 87.5% frequency among seven of IAV infected patients and increased expression of 57.15 and 27.95 fold respectively over levels of expression on the control cells. A third group with 75% frequency in six of the IAV infected patients (CCL1, IL1A, and IL1F10) increased mRNA transcriptional level 27.02,16.86 and 10.03 folds respectively. The last group showed only 50% frequency in half of the IAV infected patients with moderate to low increase in IFNA2 and IL9 mRNA expression (5.95 and 1.58 folds respectively).
Although only 21 inflammatory genes out of the 84 tested genes showed increase in their mRNA expression, six genes (CEBPB, CCR1,IL1R1, MIF, CXCL11, and IL9R) showed a lower expression than the control cells. The level of CEBPB mRNA expression was downregulated 1.37 folds in the eight IAV infected patients but with 87.5% frequency, whereas the chemokine receptor CCR1 was the most highly downregulated gene induced by IAV. The virus induced a 250 fold decrease in the expression of CCR1 transcript but with 75% frequency among the eight patients compared to its expression on control cells after IAV recovery. Cytokine IL1R1 gene expression was lower than the control with only 1.52 folds and with 75% frequency in the eight patients. The expression of CXCL1, MIF and IL9R genes showed a low decrease in their mRNA expression. The influenza A virus induced downregulation in these genes (4.93, 1.52 and 1.5 folds respectively) with a 62.5% frequency in the eight patients.
The extent of similarities and differences between the expression profiles of the 8 IAV patients were investigated using software ‘Cluster’ and displayed as heat maps using ‘Tree View’ as described previously [18]. This analysis has the advantage of demonstrating all possible relationships between the examined genes at once. Clearly, the most important component of the heat map is the last part, which shows a clear up regulation mostly in all cases. In the other heat map parts the changes were relatively insignificant and played a minor role in revealed dendrogram that segregate the 8 samples into two groups, one including the patients 1, 2, 5, 6, 7, and 8, and the other including the patients 3 and 4 (Figure 2).
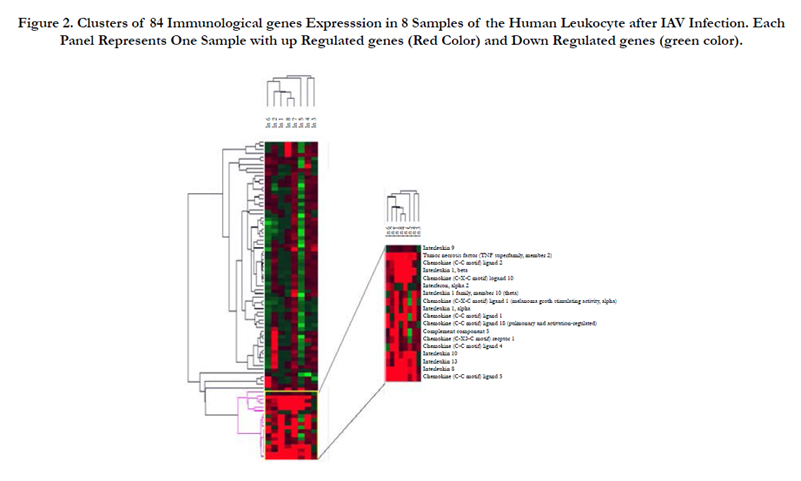

Figure 2. Clusters of 84 Immunological genes Expresssion in 8 Samples of the Human Leukocyte after IAV Infection. Each Panel Represents One Sample with up Regulated genes (Red Color) and Down Regulated genes (green color).
Discussion
The upregulation of most inducible genes in this study demonstrated early cellular response (24- 48 h), suggesting that the expression of these genes may initiate the cascades of an antiviral response. Most of upregulated genes cause asthma complication with IAV infection and play a key role in the cytokine-cytokine receptor interaction pathway [17]. The first upregulated gene group with 100% Frq includes 14 genes (Table 2, Figure 2). IL8 cytokine is a powerful leukocyte chemotactic factor that activates IL8 receptor A and IL8 receptor B. The activation of IL8 was found to lead to bronchiolitis and ends with asthma [17]. Another study showed that IL8 stimulates the innate immune system response and may cause asthma with combination to interleukins 10 [19]. In all of the eight IAV patients, IL8 cytokine was the most highly activated pro-inflammatory gene, which would indicate its involvement in the IAV inflammatory response. IL13 is known to induce B-cell differentiation and to down-regulates macrophage activity, thereby inhibiting the production of pro-inflammatory cytokines and chemokines. Thus the upregulation of IL13 transcription may be directed by IAV to reduce pro-inflammatory cytokines and chemokines. Previous studies showed that IL13 expression was found in the airways of patients with both allergic and non allergic asthma [20, 21]. Another study demonstrated that IL13 induces the pathophysiological features of asthma and is critical to allergen-induced asthma [22]. In this study IL13 cytokine was the second most highly pro-inflammatory gene product induced in all eight patients with 100% Frq. Therefore such significant upregulation of IL13 mRNA transcription after IAV infection might induce asthmatic symptoms. The β-chemokine CCL5/RANTES (regulated upon activation, normal T cell expressed and secreted) is a potent chemoattractant cytokine that recruits monocytes, memory T-cells and eosinophiles, acting as a ligand via receptors CCR1, CCR3, and CCR5 [23]. It is also able to inhibit the in vivo infection of lymphocytes [24]. CCL5 / RANTES mRNA expression was highly upregulated upon infection with IAV in this study which might be one of the factors causing asthma complicationdue to its accumulation in respiratory secretion after 24 h of IAV infectionas it has been illusterated previously [25, 23].
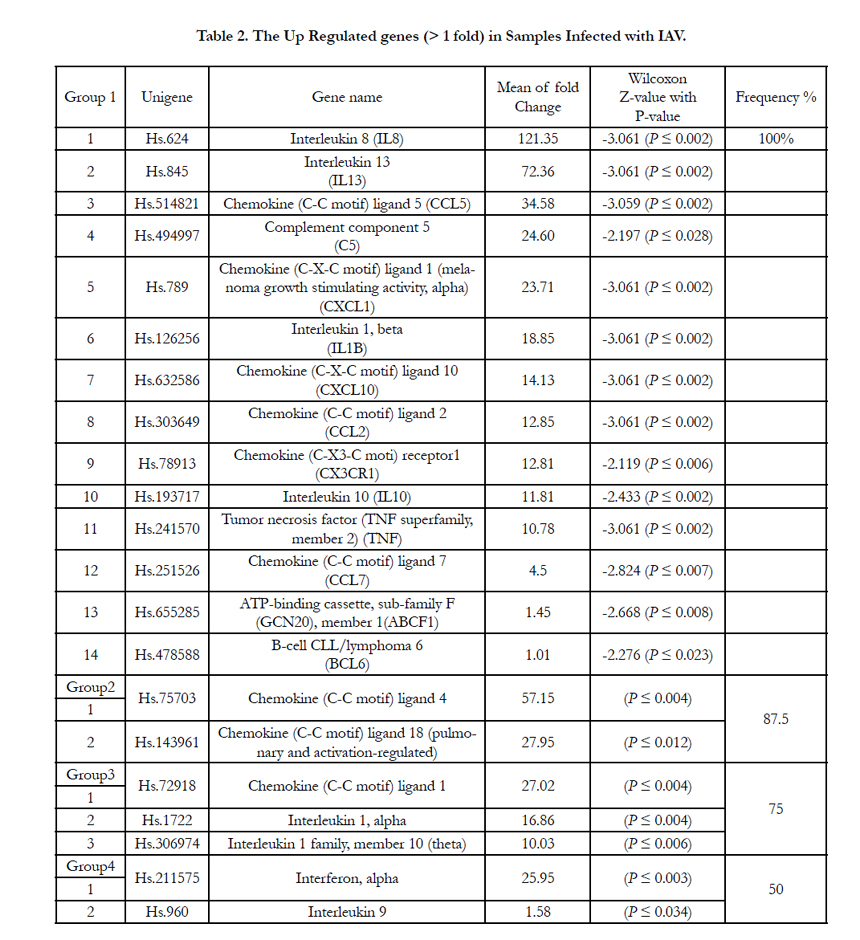
Table 2. The Up Regulated genes (> 1 fold) in Samples Infected with IAV.
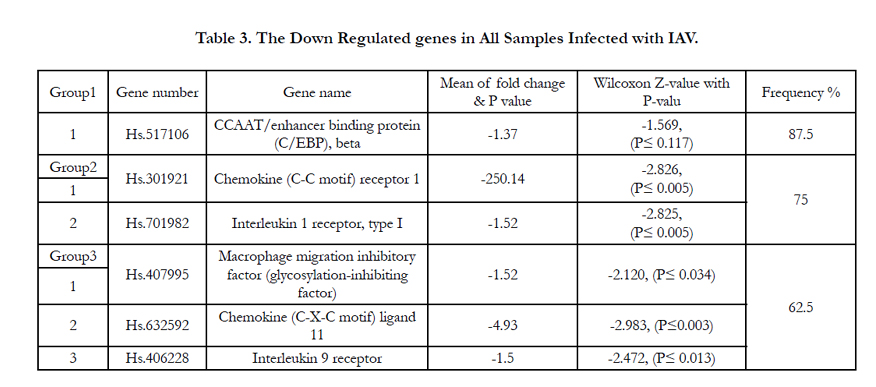
Table 3. The Down Regulated genes in All Samples Infected with IAV.
Complement-5 (C5) is a protein involved in the complement system that will be activated due to the presence of bacteria or viruses leading to cell lyses. Upon activation C5 is cleaved to C5a and C5b where C5a anaphylotoxin plays an important role in chemotaxis. This activation stimulates inflammatory responses especially in lung infections leading to smooth muscle contraction in the bronchial air ways, increases vascular permeability and causes histamine release from mast cells and basophiles [26]. Thus it causes high sensitivity which may lead to asthma [27]. C5 mRNA expression was highly upregulated in the eight IAV infected patients; this would indicate its involvement in the IAV inflammatory response. The function of the melanoma growth stimulatory activity/growth-regulated protein (CXCL1) gene is to regulate the cell trafficking of various types of leukocytes through interactions with a subset of 7-transmembrane G protein-coupled receptors. It also plays fundamental roles in the development, homeostasis, and function of the immune system [17]. In this study the CXCL1 mRNA expression is highly upregulated after 48 hrs of IAV infection which leads to the stimulation of other inflammatory genesby signal transduction activation. Previous studies reported that CXCL1 acts through G-protein to activate IL-6 gene expression since the viral infection-induced upregulation of IL-6 and IL-8 gene expression was positively correlated to the onset of acute inflammatory pain and may lead to asthma complication [28, 29]. In another study the CXCL1,showed high levels of expression (30.5-fold) during inflammation and progression of melanocytes into malignant melanoma [30]. The results of these studies are consistent with the findings of this study especially in the highly upregulation of CXCL1 after IAV infection and the presence of asthma complication signs and symptoms in these patients. IL1B cytokine is an important mediator of the inflammatory response, and is involved in a variety of cellular activities, including cell proliferation, differentiation, and apoptosis. It activates IL-1R type 2 which in turn stimulates IL-2 release, B-cell maturation and proliferation, and fibroblast growth factor activity [31]. The high significant mRNA expression of IL1B gene in this study suggests its immune role against the IAV by activation of many immune response activities which may lead to asthma symptoms. In a previous study, plasma IL1B cytokine level where detected but did not change during the course of study [32] whereas studies on variety of cells in the respiratory mucosa where found to be a source of the proinflammatory cytokines. Cultured human monocytes/ macrophages exposed to IAV produced IL1B1 in addition to IL- 6, TNF-α and IFN-α/β [33]. The CXCL10/IP-10 gene plays pleiotropic immunological effects such as stimulating monocytes, natural killer cells and T-cell migration and modulation of adhesion molecule expression [34]. In this study the IAV infection stimulates the transcription of CXCL10 mRNA at early infection with high significance which may regard as proinflammatory gene that acts early with viral infection. Others researches showed that CXCL10/IP-10 was the most upregulated gene (27-fold) in PBMCs infection [35]. Another study illustrated that CXCL10/IP-10 was secreted to induce the IFN-gamma production in lymphocytes during microbial inflammations [36]. The CCL2 has direct and indirect effect on immune responses toward viral infections through its functions as a chemotactic factor to monocytes and basophiles. It activates CCR2 which in turn acts as a receptor for the monocyte chemoattractant proteins (MCP-1, MCP2, MCP-3 and MCP-4) [16]. In the present study CCL2 transcription is upregulated significantly under the effect of IAV infection, this suggests its important immunological role to overcome the early IAV infection. These results are in agreement with previous study which showed that CCL2 is upregulated in influenza virus-infected culturesof human monocytes but with the exception that our study is in vivo whereas the previous one was in vitro [9]. CX3CR1 (chemokine fractalkine) gene has two important functions, one as a mediator for both adhesive and migratory of leukocytes to activate endothelial cells, where it is primarily expressed and second as a co-receptor for viral envelope protein [17]. In our study IAV infection stimulates the mRNA expression of this gene which may interact with IAV invasion and mediates different immunological processes. These findings are consistent with another study reported that IAV stimulates CX3CR1 mRNA to facilitate the recognition and binding with the IV-A after invading the host cell [37]. IL10 cytokine with potent anti-inflammatory effects. It repressesthe expression of TNF-α, IL6 and IL1 by activating macrophages. IL10 specific role during acute viral infection is not known yet, but a recent study reported that IL10 appears to play a detrimental role during host response to acute influenza virus infection [38]. Another study showed a significant increase in IL10 mRNA expression in virus infected subjects presenting the acute asthma and to less extent in the group with stable asthma, but not observed in virus infection with out asthma [39]. In this study IL10 mRNA expression was significantly increased after IAV infection indicating its impact on inflammatory cells, air way responsiveness and asthma occurrence. TNF gene encodes a multifunctional proinflammatory cytokines through binding to its receptors TNFRA and TNFRB leading to the translocation of transcription factor NF-kB to the nucleus which turns on the transcription of more than 60 known genes that participate in activation of proinflammatory activities and apoptosis of infected cells [40]. In this study the high transcription of TNF mRNA is significantly induced by IAV infection and its production indicates an important cellular response for viral infection in general. It was recorded previously that TNF mRNA was significantly increased by infection of influenza virus and Sendai virus [34]. Monocyte chemotactic protein 3 (MCP-CCL7) a CC chemokine that is able to attract and activate a variety of leukocytes including monocytes, natural killer cells and T-lymphocytes, by binding at least four different chemokines receptors [41]. CCL7 is implicated in several lung diseases as well as asthma, characterized by chronic inflammation and local tissue eosinophil [42]. MCP-3 (CCL7) plays an important role on monocyte/macrophage recruitment during respiratory viral infection [43, 44]. In this study, we have reported a moderate upregulation in CCL7 mRNA expression after recovery. This present data indicates that CCL7 may have an important role in the exacerbation of asthma in IAV infected patients. The ABCF1 (ATP-binding cassette/subfamily1) and BCL6 (B-cell/lymphoma 6) genes involved in the inflammatory responses were expressed in leukocytes of all IAV infected patients, but their level did not show high upregulation in upon viral infection compared to the control cells.CCL4 (MIP-1β) and CCL18 (DC-CK1/PARC/MAC-1) are one potent chemoattractants produced by NK-cells, macrophages and T-lymphocytes. They enhance the immune cell recruitment during inflammatory and viralresponses [45]. CCL18 significantly increased eosinophilic inflammation in asthma [46, 47]. In this study CCL4 and CCL18 mRNA transcripts were highly upregulated but not in all IAV infected patients, suggesting that CCL4 and/or CCL18 can be involved in this viral response but might be delayed to show their expression. This delay between the stimulus and the appearance of CCL18 has been already found in other studies [47-49]. IL1A and IL- 1F10 are members of the IL-1 family which seems to participate in the pulmonary immune response against pathogens [50]. CCL1 was found to stimulate monocyte chemotactic activity [51]. CCL1 was significantly increased in the broncheoalveolar fliud (BALF) of asthmatic patients [52]. IL1A pulmonary concentration was also elevated in H1N1 A/Puerto Rico/8/34 infected mice [53]. In our study CCL1, IL1A and IL1F10 mRNA expressions were consistent with those studies. They showed a significant upregulation in most of the IAV infected patients. Interferon alpha 2 (IFNA2) sthe most important gene for viral response which acts as; a natural cell-signaling protein produced by the cells of the immune system in response to the presence of double-stranded RNA, a key indicator of viral infection [54]. In present study the mRNA transcription of IFNA2 increased in half of the patients after 48 hrs of IAV infection. Similar results were recorded previously that IFNA2 is highly upregulated in viral infections and stimulates gene expression in NK cells and antigen processing and induces hundreds of IFN stimulated genes (ISGs) which activate the antiviral state in the infected cells [55, 56, 57]. IL9 gene is involved in inflammatory diseaseand regard as a multifunctional cytokine secreted by activated T cells and includes mastcells, B lymphocytes, T-cell clones, hematopoietic progenitors [58]. Its mRNA transcription in this study showed a low up regulation in half of the patients at least at the initial stages (48 hrs) of IAV infection. Previous study reported that IL9 show significantly higher plasma levels during viral infection [59].
Although six different inflammatory cytokines and chemokines showed a relative downregulation in their mRNA level, only CCR1 showed the most significant downregulation. The other five were almost same as control cells with little decrease. CEBPB gene is important transcriptional activator in the regulation of genes involved in immune and inflammatory responses. Specifically activate IL6 signaling pathway and plays a role in the regulation of inflammation in immune system response [16]. Thus, the down regulation of this gene may be induced by the IAV to facilitate viral replication by inhibiting the specific role of IL6 signaling pathway, therefore this gene is also called NF-IL6 gene. In another study the mRNA expression of CEBPB gene was downregulated in a combination with the effect of Ras-protein that activated under immune-inflammatory responses against microbial infection [60]. CCL4 (MIP-1β) and CCL5 (RANTES) signaling is mediated via CCR1 and CCR5. CCR1 expression and function was insufficient to induce chemokine response of human monocytes during influenza A infection with active replicating virus [61]. On the other hand, airway smooth muscle cells showed a significant level of CCR1 mRNA and protein, both in vitro and in vivo. CCR1 mRNA transcription level showed a highly significant downregulation in all the infected patients [62]. These results indicate that CCR1 expression in leukocytes is not sufficient to induce the immune response to the viral infection.
Interleukin-1α and IL1β are potent proinflammatory cytokines where their biological activity is mediated by binding to the IL- 1receptor type1 (IL1R1). Upon binding, IL1R1 recruits an accessory protein needed for its intracellular transduction [63]. IL1α expression was shown to be upregulated in most of the IAV infected patients with a 75% frequency whereas IL1R1 mRNA expression shows a slight downregulation with the same frequency (75%). These results suggest that, the reducing of IL1R1 may be a consequence at least in part of local production of IL1 at early times during IAV infection. MIF, CXCL11 and IL9 play an important role in cellular immunity in response to viral infections. On the other hand, MIF, CXCL11 and IL9 mRNA levels showed almost same levels as the control cells but with a slight decrease in their expression. This might indicate a delay in their response as been illustrated in previous studies showing that macaques infected with 1918-IAV showed a delay in CXCL11 expression. Or it indicates that they have already activated the other inflammatory cytokines to increase such as the effect of IL9 in stimulating IL8 release.
The clustering of leukocyte gene expression profile after IAV infection reveals significant alterations in the expression of inflammatory genes (Figure 2). Most patients showed upregulated genes in the lower part of the cluster. In the other heat map parts the changes were relatively insignificant and played a minor role in the dendrogram. Patients 5 and 6 showed many downregulated genes along their panels. This suggests that IAV causes viral effect on the genes of patients 5 and 6 to reduce cellular immunity responses. This general similarity in the gene expression profile suggests three points; first, the main feature of viral-leukocyte interaction is a cellular response to the viral infection, second, all patients are mostly infected with related subtypes of influenza A viruses, third, the patients 1, 2, 6, 7 and 8 are related since they originated from one cluster, whereas 3 and 4 are more similar andclustered from one origin.
Conclusion
Our results reveal alterations in host inflammatory gene expression induced by influenza A virus. The over expressed genes with 100% Frq represent the leukocyte responses to IAV. Eight of upregulated genes are involved in asthma complication in patients with IAV. The downregulated genes represent the viral effect on leukocyte. All patients are mostly infected with related subtypes of influenza A viruses. These findings may provide new targets for further analysis of influenza A virus role in the viral-leukocyte interaction.
Acknowledgements
This work is supported by grants from the University of Jordan, Scientific research council, faculty of graduate studies. We thank Dr. Malek Zihlif for cooperation in genes analysis; Dr Maher Taka for help in Statistical analysis; all staff of Molecular Biology Lab, Medicine College, University of Jordan for their help during the period of practical work.
References
- Eccles R (2005) Understanding the symptoms of the common cold and influenza. Lancet Infect Dis. 5(11): 718–725.
- Bresson J, Christian P, Odile L, Catherine G, Melanie S,et al., (2006) Safety and immunogenicity of an inactivated split-virion influenza A/Vietnam/1194/2004 (H5N1) vaccine: phase I randomised trial. Lancet.3670:1657-1664.
- Treanor JJ (1999) Influenza A and B viruses: Viral Infections of the Respiratory Tract; (2nded). New York: Marcel Dekker, 1999, 105–160.
- See H, Wark P (2008) Innate immune response to viral infection of the lungs. Paediatr Respir Rev. 9(4): 243–250.
- David F, Farley J, Huang H, Lavoie JP, Laverty S (2008) Cytokine and chemokine gene expression of IL-1beta stimulated equine articular chondrocytes. Vet Sur. 36(3): 221-227.
- Melchjorsen J, Sorensen LN, Paludan SR (2003) Expression and function of chemokines during viral infections: from molecular mechanisms to in vivo function. J Leukoc Biol. 74(3): 331-343.
- Geiss GK, Salvatore M, Tumpey TM (2002) Cellular transcriptional profiling in influenza A virus-infected lung epithelial cells: the role of the nonstructural NS1 protein in the evasion of the host innate defense and its potential contribution to pandemic influenza. Proc Natl Acad Sci USA. 99:10736–10741.
- Bergmann M, Adolfo G, Elena C, Hubert P, Klaus W, et al., (2000) Influenza Virus NS1 Protein Counteracts PKR-Mediated Inhibition of Replication. J Virol. 74(13): 6203-6206.
- Bubfeld D, Kaufmann A, Meyer RG, Gemsa D, Sprenger H (1998) Differential mononuclear leukocyte attracting chemokine production after stimulation with active and inactivated influenza A virus. Cell Immunol.186(1):1–7.
- Sprenger H, Meyer RG, Kaufmann A, Bubfeld D, Rischkowsky E, et al., (1996) Selective Induction of monocyte and not nutrophile attracting chemokines after influenza A virus Infection. J Exp Med. 184(3): 1191-1196.
- Tong Hua Hua, James P Long, Daneng Li, Thomas F (2004) DeMaria: Alteration of gene expression in human middle ear epithelial cells induced by influenza A virus and its implication for the pathogenesis of otitis media.Microbial pathogenesis. 37(4): 193-204.
- Tombacz D, Toth JS, Petrovszki P, Boldogkoi Z (2009) Whole-genome analysis of pseudorabies virus gene expression by real-time quantitative RTPCR assay. BMC Genomics.10: 491.
- Myskiw C, Arsenio J, Van BR, Deschambault Y, Cao J (2009) Vaccinia virus E3 suppresses expression of diverse cytokines through inhibition of the PKR, NF-KappaB, and IRF3 pathways. J Virol. 83(13): 6757 -67568.
- Riny J, Jeroen P, Hennie H, Annemarie B, Marijike van O, et al., (2005) Host transcription profiles upon primary respiratory syncytial virus infection. J Virol. 81(11): 5958-5967.
- Sambrook J, Fritsch EF, Maniatis T (1989) Molecular Cloning: A Laboratory manual. (3rd Edn), Cold Spring Harbor Laboratory, Cold Spring Harbor, New York.
- Joshua SY, Ann R, Feng C, Neal Jr (2006) Statistical analysis of real-time PCR data. BMC Bioinformatics. 7(1): 1.
- Huang DW, Sherman BT, Lempicki RA (2009) Systematic and integrative analysis of large gene lists using DAVID Bioinformatics Resources. Nat Protoc. 4(1): 44-57.
- Eisen MB, Spellman PT, Brown PO, Botstein D (1998) Cluster analysis and display of genome-wide expression patterns. Proc Natl Acad Sci U S A. 95(25): 14863-14868.
- Utgaard JO, Jahnsen FL, Bakka A, Brandtzaeg P, (1998) Rapid secresion of prestord interleukin 8 from Wwibel-Palade bobies of microvascular endothelial cells. J Exp Med. 188(9): 1751–1756.
- Walker C, Bode E, Boer L, Hansel TT, Blaser K, et al., (1992) Allergic and nonallergic asthmatic have distinct patterns of T-cell activation and cytokine production in peripheral blood and bronchoalveolar lavage. Am Rev Respir Dis.146(1):109-115.
- Humbert M, Durham S, Kimmitt P, Powell N, Assoufi B, et al., (1997) Elevated expression of messenger ribonucleic acid encoding IL-13 in thebronchial mucosa of atopic and nonatopic subjects with asthma. J Allergy Clin Immunol. 99(5): 657-665.
- Wills-Karp M, Luyimbazi J, Xu X, Schofield B, Neben TY, et al., (1998) Interleukin- 13: central mediator of allergic asthma. Science. 282: 2258-2261.
- Cully FJ, Alasdair MJ, Pennycook JS, Tregoning JS, Jonathan SD, et al., (2006) Role of CCL5 (RANTES) in viral lung disease. J Virol. 80(16): 8151-8157.
- Cocchi F, DeVicoAL, Garzino-DimoA, Arya SK, Gallo RC, (1995) Identification of RANTES, MIP-1 alpha and MIP-1 beta as the major HIV-suppressive factors produced by CD8+ cells. Science. 270(5243): 1811-1815.
- Brydon A, Harry S, Clive S (2003) Influenza A virus-induced apoptosis in bronchiolar epithelial (NCI-H292) cells limits pro-inflammatory cytokine release. J Gen Virol. 84(9): 2389-2400.
- Shushakova N, Sokowa J, Schulman J, Baumann U, Zwirner J, et al., (2002) C5a anaphylatoxin is a major regulator of activating versus inhibitory FcgammaRs in immune complex-induced lung disease. J Clin Invest.110(12):1823-1830.
- Qian FH, Zhange Q, Zhou LF, Liu H, Huang XL, et al., (2008) Highsensitivity C-reactive protein: a predictive marker in severe asthma. Resoirology. 13(5): 664-669.
- Papdopoulos NG, Papi A, Meyer J, Stanciu LA, Salni S, et al., (2001) Rhinovirus infection up-regulatseotaxin and eotaxin -2 expression in bronchial epithelial cells. Clin Exp Allergy. 31(7): 1060-1066.
- Wang XM, Hamza M, Wu TX, Dionne RA (2009) Upregulation of IL-6, IL-8 and CCL2 gene expression after acute inflammation: Correlation to clinical pain. Pain. 142(3): 275-283.
- Crompton MR, Moss SE, Crumpton MJ (1988) Diversity in the lipocortin/ calpactin family. Cell. 55: 1–3.
- Stuart Allan, Nancy Rothwell (2001) Cytokines and Acute Neurodegeneration. Nat Rev Neurosci. 2(10): 734-744.
- Hayden FG, Fritz R, Lobo MC, Trober W, Straus SE (1998) Local and systemic cytokine responses during experimental human influenza A virus infection relation to symptom formation and host defense. J Clin Invest. 101: 643-649.
- Peschke T, Bender A, NainM, Gemsa D (1993) Role of macrophage cytokines in influenza A virus Infections. Immunobiology.189(3-4): 340-355.
- Veckman Ville, Pamela Osterlund, RikuFagerlund, KristerMelen, SampsaMatikainen et al., (2006) TNF-α and IFN-α enhance influenza-A-virus -induced chemokine gene expression in human A549 lung epithelial cells.Virology. 345(1): 96-104.
- Hutchinson KL, Villinger F, Miranda ME, Ksiazek TG, Peters CJ, et al., (2001) Multiplex analysis of cytokines in the blood of cynomolgus macaques naturally infected with Ebola virus (Reston serotype). J Med Virol. 65(3):561–566.
- Dufour JH, Dziejman M, Liu MT, Leung JH, Lane TE, et al., (2002) IFN gamma-inducible protein 10 (IP-10 CXCL10)-deficient mice reveal a role for IP-10 in effect on T cell generation and trafficking. J Immunol. 168(7):3195–3204.
- Matloubian M, David A, Engel S, Ryan JE, Cyster JG (2000) A transmembrane CXC chemoline is aligant for HIV-coreceptorBonzo. Nat Immunol. 1(4): 298-304.
- Keer Sun, Luisa Torres, Dennis W Metzger (2008) A Detrimental Role for IL-10 during Host Immune Responses to Influenza Virus Infection.FASEB J. 22: 857-865.
- Grissell TV, Powell H, Shafren DR, Boyle MJ, Michael J, et al., (2005) Interleukin-10 Gene Expression in Acute Virus-induced Asthma. Am J Resp Crit Care Med. 172(4): 433-439.
- Mehta DS, Wurster AL, Whitters MJ, Young DA, Collins M, et al., (2003)IL12 induces the apoptosis of resting and activated primary B cells. J Immunol.170(8): 4111-4118.
- Menten Patritia, Ania Wuyts, Jo Van Damme (2001) Monocyte chemotactic protein-3. European cytokine network network. 12(4): 554-560.
- MiottoDeborah ,PotaChristodoulopoulos , Ron Olivenstein , RameTaha , Lisa Cameron, et al., (2001) Expression of IFN-γ–inducible protein; monocyte chemotactic proteins 1, 3, and 4; and eotaxin in TH1- and TH2-mediated lung diseases, J Allergy Clin. Immunol. 107(4): 664-670.
- Rojas-Ramos E, AF Avalos, L Perez-Fernandez, F Cuevas-Schacht, E Valencia- Maqueda et al., (2003) Role of the chemokines RANTES, monocyte chemotactic proteins-3 and -4, and eotaxins-1 and -2 in childhood asthma. EurRespir J. 22(22): 310-316.
- Santiago J, JL Hernandez-Cruz, ME Manjarrez-Zavala, R Montes-Vizuet, DP Rosete- Olvera, et al., (2008) Role of monocyte chemotactic protein- 3 and -4 in children with virus exacerbation of asthma. Eur Respir J. 32(5):1243-1249.
- Guan E, Wang J, Roderiquez G, Norcross MA (2002) Natural truncation of the chemokine MIP-1 beta /CCL4 affects receptor specificity but not anti- HIV-1 activity. J Biol Chem. 277(35): 32348-32352.
- Kim HB, Kim CK, Iijima K, Kobayashi T, Kita H (2009) Protein microarray analysis in patients with asthma: elevation of the chemokine PARC/ CCL18 in sputum. Chest. 135(2): 295-302.
- De Nadai Patricia, Anne-Sophie Charbonnier, Cecile Chenivesse, Stephanie Senechal, Clement Fournier, et al., (2006) Involvement of CCL18 in Allergic Asthma. J Immunol. 176(10): 6286-6293.
- KodeljaVitam, Carola Müller, Oliver Politz, NahidHakij, Constantin E et al., (1998) Alternative Macrophage Activation-Associated CC-Chemokine-1, a Novel Structural Homologue of Macrophage Inflammatory Protein-1 with a Th2-Associated Expression Pattern. J Immunol.160(3): 1411-1418.
- Goebeler Matthias, Axel Trautmann, Ariane Voss, Eva-Bettina Brocker, AtiyeToksoy, et al., (2001) Differential and Sequential Expression of Multiple Chemokines during Elicitation of Allergic Contact Hypersensitivity. Am J Pathol. 158(2): 431-440.
- Rosseau Simone, Andreas Hocke, Hans Mollenkopf, Bernd Schmeck, Norbert Suttorp, et al., (2007) Comparative transcriptional profiling of the lung reveals shared and distinct features of Streptococcus pneumoniae and influenza A virus infection Immunology. 120(3): 380–391.
- Oliveira SH, Lira S, Martinez-A C, Wiekowski M, Sullivan Let al., (2002) Increased responsiveness of murine eosinophils to MIP-1beta (CCL4) and TCA-3 (CCL1) is mediated by their specific receptors, CCR5 and CCR8. J LeukocBiol. 71(6):1019–1025.
- Montes-Vizuet R, Vega-Miranda A, Valencia-Maqueda E, Negrete-Garcia MC, Velasquez JR, Et al., (2006) CC chemokine ligand 1 is released into the airways of atopic asthmatics. Eur Respir J 2006; 28(1):59-67.
- Aleksandr S. Lipatov, SamitaAndreansky, Richard J. Webby, Diane J. Hulse, Jerold E. Rehg, et al., (2005) Pathogenesis of Hong Kong H5N1 influenza virus NS gene reassortants in mice: the role of cytokines and B- and T-cell responses. J Gen Virol. 86(4):1121–1130.
- Kotenko SV, Gallagher G, Baurin V, Lewis-Antes A, Shen M, et al., (2003) IFN-lambdas mediate antiviral protection through a distinct class II cytokine receptor complex. Nat Immunol. 4(1): 69-77.
- Matikainen S, Paananen A, Miettinen M, Kurimoto M, Timonen T, et al., (2001) IFN-alpha and IL-18 synergistically enhance IFN-gamma production in human NK cells: differential regulation of Stat4 activation and IFNgamma gene expression by IFN-alpha and IL-12. Eur J Immunol. 31(7):2236-45.
- Dalod M, Salazar-Mather TP, Malmgaard L, Lewis C, Asselin-Paturel C, et al., (2002) Interferon alpha/beta and interleukin 12 responses to viral infections: pathways regulating dendritic cell cytokine expression in vivo. J Exp Med. 195(4): 517-528.
- Sato M, Suemori H, Hata N (2005) Distinct and essential roles of transcription factors IRF-3 and IRF- 7 in response to viruses for IFN alpha/beta gene induction. Immunity.13(4): 359-548.
- Ryan AW, Thornton JM, Brophy K, Daly JS, Mcloughlin RM, et al., (2005) Chromosome 5q candidate genes in coeliac disease: genetic variation at IL4, IL5, IL9, IL13, IL17B and NR3C1. Tissue Antigens. 65(2):150-5.
- Klingstrom Jonas, Therese Lindgren, ClasAhlm (2008) Hantavirus infection induces higher plasma levels of IL-9, FGF-2 and Gm-CSF, and lower levels of IL-8 and IP-10, in females compared to males,Vaccine Immunol.doi:10.1128/CVI.00035-08.
- Sebastian T1, Johnson PF (2009) RasV12-Mediated Down-regulation of CCAAT/Enhancer Binding Protein β in Immortalized Fibroblasts Requires Loss of p19Arf and Facilitates Bypass of Oncogene-Induced Senescence.Cancer Research. 69(6): 2588-98.
- Schmitz N, Kurrer M, Bachmann MF, Kopf M (2005) Interleukin-1 Is Responsible for Acute Lung Immunopathology but Increases Survival of Respiratory Influenza Virus Infection Journal of Virology. 79(10): 6441-6448.
- Zhu W, Higgs B, Morehouse C, Streicher K, Ambrose C, et al., (2010) A whole genome transcriptional analysis of the early immune response induced by live attenuated and inactivated influenza vaccines in young children, Vaccine.28(16): 2865–2876.
- Zlotnik A, Yoshie O, Nomiyama H, (2006) The chemokine and chemokine receptor superfamilies and their molecular evolution, Genome Biology. 7(1):243.





