Side Population Cells in Human Adenomyotic Lesions Display Characteristics of Endometrial Stem Cells
Zhu S1, Xiao L1, Huang W1*, Zhou M1, Zhang X1, Yang S1, Xu W2, Sun H2
1 Department of Obstetrics and Gynecology, West China Second University Hospital of Sichuan University, Chengdu Sichuan, P. R. China.
2 Sichuan University - The Chinese University of Hong Kong Joint Laboratory for Reproductive Medcine, West China Institute of Women and Children’ Health, West China Second University Hospital, Sichuan University, Chengdu Sichuan P.R. China.
*Corresponding Author
Wei Huang,
Department of Obstetrics and Gynecology,
West China Second University
Hospital of Sichuan University, Chengdu Sichuan 610041,
P. R. China.
Tel: +8613881858999
Email: weihuang64@163.com
Article Type: Research Article
Received: April 02, 2015; Accepted: May 12, 2015; Published: May 14, 2015
Citation: Huang W, et al., (2015) Side Population Cells in Human Adenomyotic Lesions Display Characteristics of Endometrial Stem Cells. Int J Stem Cell Res Transplant, S1:001, 1-6. doi: dx.doi.org/10.19070/2328-3548-SI01001
Copyright: Huang W© 2015. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
Introduction: The precise etiology and physiopathology of ademyosis is still unknown. The aim of this research is to investigateif stem cells are found in adenomyosis and to characterize the role of adenomyotic lesion-derived side population (SP) cells in pathogenesis of adenomyosis.
Methods: We recruited 31 women (range 37-48 years) treated by hysterectomy for adenomyosis. Adenomyotic samples were collected and The SP cells in the tissues were identified by flow cytometry. ATP-binding cassette transporter 2+ (ABCG2+ ) cells were isolated from human adenomyotic samples by magnetic activated cell sorting (MACS), further subjected to colony formation, induced differentiation in four conditions and mobility/invasion assay. The cell lines generated from ABCG2+ cells were analyzed by immunofluorescence, quantitative real-time PCR and western blot analysis.
Results: The side population represents 1.59±0.48% of the total cell population in adenomyotic lesions. Cloning efficiency of the ABCG2+ cells was 11.27±1.74%. When co-cultured with endometrial cells, ABCG2+ cells differentiated into endometrium-like cells. Moreover, if 17β-estradiol was administrated to the co-culture system, a significantly higher efficiency of differentiation was detected (P<0.05). ABCG2+ cells in co-culture system displayed a higher efficiency of differentiation, compared to that in culture without feeder cells (p<005). The wound closure and transwell assays demonstrated that 17β-estradiol stimulates the migration/invasion of ABCG2+ cells in a dose-dependent manner, with a peak effect at a concentration of 10-8 M.
Conclusions: The adenomyosis-derived ABCG2+ cells display stem cell-like properties and may be involved in the pathogenesis of adenomyosis.
2.Introduction
3.Materials and Methods
3.1.Human tissue samples
3.2.Hoechst 33342 labeling for flow cytometry
3.3.Isolation of ABCG2+ cells, cell culture and colony-forming
3.4.Preparation of feeder cells
3.5.In vitro induced differentiation
3.6.Immunofluorescence
3.7.Western blot analysis
3.8.Quantitative real-time PCR
3.9.Migration and invasion assay
3.9.Statistical analysis
4.Results and Discussion
4.1.Identification of SP cells from human adenomyotic tissues
4.2.Clonogenicity of adenomyotic lesions-derived ABCG2+ cells
4.3.Induced differentiation of ABCG2+ cells from adenomyotic lesions
4.4.17β-estradiol enhances migration and invasion in ABCG2+ Cells
5.Conclusion
6.Acknowledgement and Declarations
7.References
Keywords
Adenomyosis; Stem Cell; Side Population Cell; Differentiation.
Introduction
Adenomyosis is a common gynecologic disorder characterized by the growth and invasion of endometrial tissue into the myometrium. It preferentially affects multiparous women in their reproductive or perimenopausal years, ranging from 14% to 66% in hysterectomy specimens [1]. However, the precise etiology and physiopathology of adenomyosis is still unclear. Several studies have been taken to form the explanations. It is considered that adenomyosis result of trauma, either mechanical or physiologic peristaltic processes [2]. Someone suggested a more conspicuous presentation based on hypervascularity and the presence of abnormal endometrial proliferation in adenomyotic lesions [3]. In addition, hormonal mediation, particularly estrogen secretion, has long been postulated as an underlying mediator [4]. Locally high level of estrogen may play an important role in endometrium invasion since high frequency of endometrial hyperplasia is found in women with adenomyosis [5]. Estrogen is thus closely associated with the growth and development of adenomyosis. However, the molecular mechanism estrogen triggers adenomyosis is unclear and requires further research.
Stem cells are undifferentiated cells that are defined by their ability to self-renew and differentiate into mature cells. Side -population (SP) cells, referred to as stem-like cell subpopulations, have been identified based on their unique ability to pump out intracellular DNA-binding dye Hoechst 33342 via ABCG2 [6]. Recently, SP cells were identified and isolated in the basalis layer of endometrium. These SP cells functioned as progenitor cells, showing longterm proliferating capacity in cultures and producing both gland and stromal-like cells [7, 8]. Furthermore, SP cells associated with abnormal endometrial proliferation have been implicated in the pathology of endometrial disease, including endometriosis and endometrial carcinoma [9, 10]. Those studies on stem/progenitor cells add new perspectives to the pathogenesis of adenomyosis. Adenomyosis is characterized by abnormal endometrial cell growth inside myometrium. It is possible that putative endometrial stem cells may play a role in the pathophysiology of adenomyosis [11]. To the best of our knowledge, few data are currently available on this issue, and it would be interesting to perform additional studies to confirm the hypothesis.
To date, SP cells have been isolated from various adult tissues and extensively characterized, implicating the SP phenotype as a common feature of adult stem cells [12]. Additionally, ABCG2 was considered as a molecular determinant of the SP phenotype in tumor stem-like cells [13]. Together, these findings indicate that ABCG2 expression is a conserved feature of stem cells from a wide variety of sources. Therefore, ABCG2 is an attractive candidate marker for identifying and isolating stem cells.
In the present study, we identified and isolated SP cells from adenomyotic tissues via ABCG2 antibodies and evaluated their characteristics in terms of colony-forming capacity, induced differentiation, migration and invasion. Subsequently, we tested the effect of 17β-estradiol, as a possible tool to modulate induced differentiation and migration/invasion properties in SP cells from adenomyotic lesions.
Materials and Methods
The study was approved by the institutional ethics board of West China Second University Hospital of Sichuan University and carried out according to the principles of the Helsinki Convention. Written informed consent was obtained from each participant.
Adenomyotic samples were obtained from 31 women (range 37-48 years, mean age 42.4±3 years) underwent hysterectomy for adenomyosis in the West China Second Hospital of Sichuan University from March 2012 to July 2013. Typical adenomyotic lesions were significantly thickening and hardening without clear boundaries in uterine wall, composed of microcapsules and non-striped muscle bundles running in all directions. The obvious adenomyotic tissue was collected and transferred to the laboratory in Dulbecco modified Eagle medium/Hams F-12 (DMEM/F-12, GIBCO, New York, USA). For feeder cells in induced differentiation, normal endometrial tissues were collected from women (range 22-34 years, mean age 26.8±3 years) with benign teratoma of ovary, but no evidence of endometriosis or adenomyosis during surgery. Patients with endometrial disorders, gynecological malignancies and precancerous lesions were excluded. None of the patients had received any hormonal treatment within 3 months.
The adenomyotic tissues were cut up manually into small pieces of < 1mm3 and incubated for 4-7 h in DMEM/F-12 containing 0.2%(wt/vol) collagenase III (Sigma, St. Louis, USA) and 1% penicillin/streptomycin at 37°C on a shaker. We removed undigested
tissues by using a 400- μm strainer (BD–Falcon, New Jersey, USA) and obtained single-cell suspensions by a 10- μm filter.
Cells isolated from the adenomyotic tissue were resuspended at a concentration of 2×106 cells/ml in calcium- and magnesium- free Hanks balanced salt solution containing 2% fetal bovine serum (FBS; GIBCO, New York, USA) and 1% penicillin/streptomycin. The cell suspension was labeled with 5μg/ml of Hoechst 33342 dye or in combination with 100μM verapamil (Sigma-Aldrich, Missouri, USA) in a water bath at 37°C for 90min. Then, SP cells were identified by flow cytometry (BD Biosciences, New Jersey, USA).
ABCG2+ cells suspensions were obtained by positive selection using MACS ( Miltenyi Biotec, Cologne, Germany) coated with antibody against ABCG2 (Abcam, Eugene, Oregon, USA) according to manufacturer’s instructions. Isolated ABCG2+ cells were seeded in triplicate on 60-mm Petri dishes at a density of 10-20 cells/cm2. The culture medium is DMEM/F-12 containing 10% FBS with stem cell growth factor and vascular endothelial growth factor (TEBU, Frankfurt, Germany) at 10ng/ml final concentration. ABCG2+ cells were cultured for 21 days. Clusters of cells were considered colonies when they were visible macroscopically and contained greater than 50 cells. Colonies were counted and the cloning efficiency (CE) determined from the formula CE (%) = (number of colonies/number of cells seeded)×100%.
The endometrial tissues were cut up manually into small pieces of < 1mm3 and incubated for 1 h in DMEM/F-12 containing 0.2% (wt/vol) collagenase III (Sigma, St. Louis, USA) and 1% penicillin/ streptomycin at 37°C on a shaker. Undigested tissues were removed by using a 400- μm strainer. The endometrial cells were seeded in six-well plates. Then mitomycin C solution was added to culture hole containing 80-90% confluent endometrial cells to achieve a 20ug/ml final concentration. After incubating for 1 h at 37°C in humidified incubator with 5% CO2, culture medium containing mitomycin C was moved.
To further characterize isolated ABCG2+ cells, we cultivated them on chamber slides using DMEM/F12/10% FBS with various additional conditions: 10-8M 17β-estradiol (Sigma-Aldrich, St. Louis, USA) with or without endometrial cells (17β-estradiol+ endometrial cells+, 17β-estradiol+ endometrial cells-), or endometrial cells alone (17β-estradiol- endometrial cells+); ABCG2+ cells cultured in DMEM/F12/10% FBS without 17β-estradiol or endometrial cells (17β-estradiol- endometrial cells-) served as a control. When co-cultured with endometrial cells, ABCG2+ cells on chamber slides were placed over them. After 3 weeks, cells on the slides in four different conditions were further subjected to immunofluorescence, qRT-PCR, and western blot analysis.
After 3-week of induced differentiation, generated cells on chamber slides were fixed in 4% paraformaldehyde for 15min, permeabilized with 0.2% Triton X-100 (Sigma) for 8min at room temperature. After blocking with 10% FBS for 8 min, slides were incubated overnight at 4°C with mouse anti-cytokeratin (1:400, Abcam, Eugene, Oregon, USA) or rabbit anti-Vimentin (1:500, Abcam, Eugene, Oregon, USA) antibodies. Primary antibodies were detected by incubation with corresponding IgG secondary antibodies conjugated with Alexa Fluor 594 (red) and 488 (green) (1:500, Invitrogen, Eugene, Oregon, USA) for 1 hr. Nuclei of the cells were counterstained with 6-diamino-2-phenylindole (DAPI; Sigma-Aldrich, USA). Substitution with an unrelated rabbit or mouse IgG served as negative control.
Cells on the chamber slides were washed twice with ice-cold phosphate- buffered saline and lysed in RIPA buffer (20 nM Tris-Hcl, 150 nM NaCl, 1% deoxycholate, 0.1% sodium dodecyl sulfate, and 1% Triton X-100, pH 7.8) for 30 min on ice. Equal amounts of protein (20μg) were separated by electrophoresis on a 10% SDS–PAGE and transferred onto a PVDF membrane. The membrane was blocked with 5% non-fat dry milk in Tris-buffered saline with Tween 20 for 1 h and incubated overnight with the primary antibodies (1:5000, mouse anti-cytokeratin or 1:10000, rabbit anti- Vimentin, separately). The bound antibodies were detected using the appropriate peroxidase-coupled secondary antibodies and visualized using a chemiluminescence detection system (Thermo). β-actin (1:3000, Kangchen Biotech Corporation, Shanghai, China) were used as protein load control.
After 3-week induced differentiation, total RNA was isolated from generated cells on chamber slides using Trizol with genomic DNA removed. A 1 μg aliquot was reverse transcribed into cDNA with RTase and random hexamers from Ex-Script reagent kit (TAKARA, Dalian, China) according to manufacturer’s instructions. Quantitative RT-PCR was performed using a Bio-Rad CFX96 system (Bio-Rad, USA) and iTaq™ SYBR green supermix (Bio- Rad) and the following specific gene primers: for human cytokeratin, 5’-AATGGGAGGCATCCAGAACGAGAA-3’ (forward) and 5‘-TTCTTCTCCAAGTGCTCCCGGATT-3’ (reverse); for human Vimentin, 5’-GGAGAAATTGCAGGAGGAGATG-3‘ (forward) and 5 ‘-AAGGTCAAGACGTGCCAGAGA-3‘ (reverse); and Gene expression levels were normalized to that of the housekeeping gene human GAPDH 5‘-CTGACTTCAACAGCGACACC-3’ (forward) and 5’ -CCCTGTTGCTGTAGCCAAAT-3’ (reverse) .
The sorted ABCG2+ cells were plated in 60-mm dishes at equivalent cell densities for the wound closure assays in DMEM/F-12 with 17β-estradiol at a final concentration gradient as 0, 10-9, 10-8, and 10-7 M. when the cells reached 80–90% confluence, a linear wound was incised in the central area of the monolayers by a pipette tip. Wound healing was observed at 0 and 48h within the scrape line, and the gap was measured.
The transwell assays were performed using 8-μm pore transwell chambers in 24-well plates (Millipore, Switzerland). The ABCG2+ cells were pretreated in DMEM/F-12 medium with 17 β-estradiol at a final concentration gradient as 0, 10-9, 10-8 and 10-7 M for 24 h. Then the cells (1×105 cells in 50ml of suspension) were plated in the upper chamber with DMEM/F-12 medium containing 0.5% FBS and parallel medium containing 20% FBS was added to the lower chamber.
For invasion assay, the upper chamber was coated with 1:5 Matrigel (BD Bioscience, New Jersey, USA). After 36h of incubation, the cells that had invaded to the lower side of the membrane were fixed with methanol, and stained with crystal violet solution. 5-8 unit fields per membrane were counted at 200×magnification using a Zeiss microscope (Carl Zeiss, New York, USA).
All experiments were performed in triplicate and repeated 3 times independently. The GraphPad Prism 3.00 was used for the statistical analyses. All the data are normally-distributed and expressed as the mean±SD. Comparisons between two groups were performed by Student’s t test, and differences among multiple groups were evaluated by one-way ANOVA. A P value of < 0.05 was considered to be statistically significant.
Results and Discussion
SP cells were isolated from 17 human adenomyotic specimens and represented 1.59±0.48% of the total cell population (Figure. 1A, left panel), but disappeared upon treated with 100μM Verapamil (Figure. 1A, right panel). We successfully isolated ABCG2+ cells in adenomyotic lesions from 17 samples as a follow-up to our previous protocol.
ABCG2+ cells displayed their stem-like properties in clone-forming (Figure. 1B). In the first week, the growth rate for the colony was very slow, with colonies generally comprising less than 100 cells on day 7. However, a majority of colonies increased dramatically from day 10 to 15. During 16-21 days, the colonies were slowly increasing in size. On day 21, large colonies contained as many as 1000 cells can be seen. The cloning efficiency of the isolated ABCG2+ cells was 11.27±1.74%.
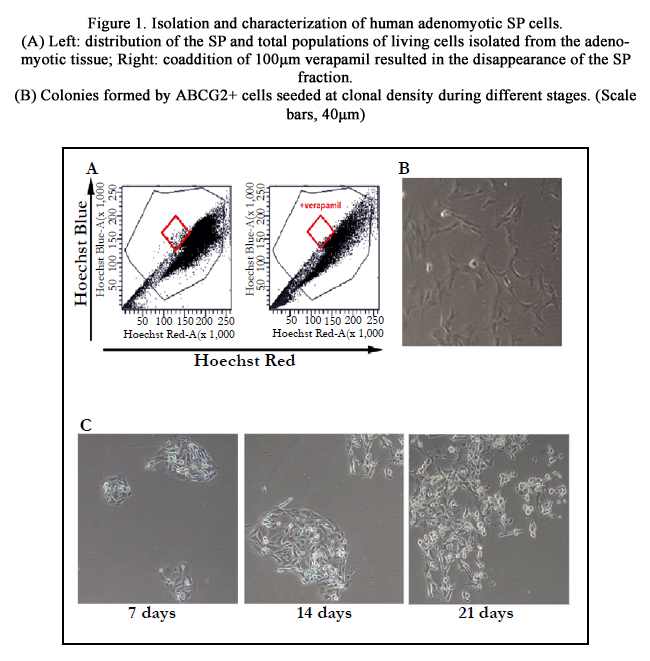
Figure 1. Isolation and characterization of human adenomyotic SP cells.
(A) Left: distribution of the SP and total populations of living cells isolated from the adenomyotic tissue; Right: coaddition of 100μm verapamil resulted in the disappearance of the SP fraction.
(B) Colonies formed by ABCG2+ cells seeded at clonal density during different stages. (Scale bars, 40μm)
Immunofluorescence analyses detected that the newly generated cells expressed protein Vimentin and Cytokeratin after ABCG2+ cells had been cultured in induction medium for 3 weeks, suggesting the potential of ABCG2+ cells for differentiation into endometrium-like cells. Moreover, the stromal cell marker Vimentin accounted for a higher proportion than epithelial cell marker Cytokeratin (Figure. 2). Differentiation efficiency of ABCG2+ cells in the four different induction mediums analyzed by western blot and qRT-PCR was shown in Figure. 3. Compared with ABCG2+ cells cultured in conditions without endometrial cells, that co-cultured with the same feeder cells displayed significantly higher differentiation efficiency (P<0.01). No difference has been observed for the Vimentin and Cytokeratin expression between culture with 17β -estradiol- endometrial cells- and 17β-estradiol+ endometrial cells- (P >0.05). However, when ABCG2+ cells were cultured in condition with 17β -estradiol+ endometrial cells+, Vimentin and Cytokeratin exhibited higher expression than that in condition with 17 β-estradiol- endometrial cells+ (P<0.01). Additionally, the expression level of cytokeratin and Vimentin determined by western blotting and qRT-PCR was consistent with the immunofluorescence analyses (Figure. 2 and 3).
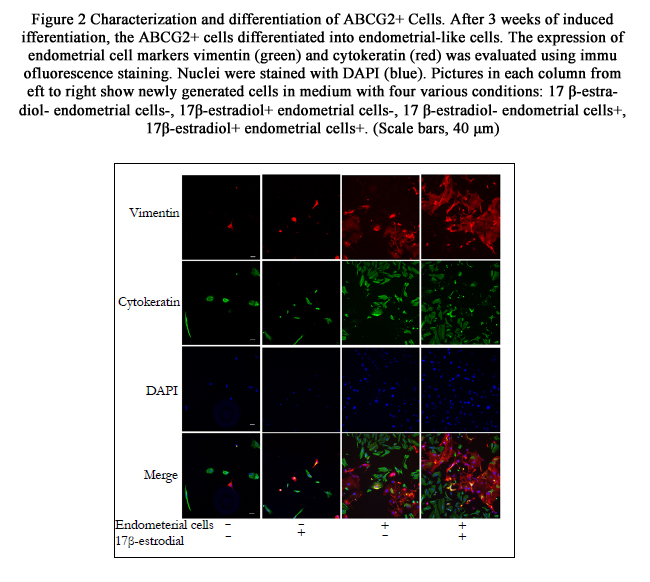
Figure 2 Characterization and differentiation of ABCG2+ Cells.
After 3 weeks of induced differentiation, the ABCG2+ cells differentiated into endometrial-like cells. The expression of endometrial cell markers vimentin (green) and cytokeratin (red) was evaluated using immunofluorescence staining. Nuclei were stained with DAPI (blue). Pictures in each column from left to right show newly generated cells in medium with four various conditions: 17 β-estradiol- endometrial cells-, 17β-estradiol+ endometrial cells-, 17 β-estradiol- endometrial cells+, 17β-estradiol+ endometrial cells+. (Scale bars, 40 μm)
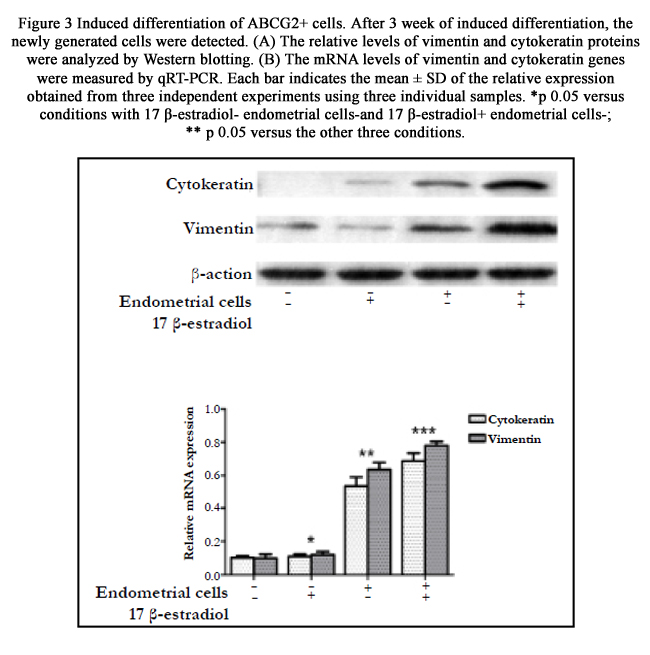
Figure 3 Induced differentiation of ABCG2+ cells. After 3 week of induced differentiation, the newly generated cells were detected.
(A) The relative levels of vimentin and cytokeratin proteins were analyzed by Western blotting.
(B) The mRNA levels of vimentin and cytokeratin genes were measured by qRT-PCR. Each bar indicates the mean ± SD of the relative expression obtained from three independent experiments using three individual samples. *p﹤0.05 versus conditions with 17 β-estradiol- endometrial cells-and 17 β-estradiol+ endometrial cells-;
** p﹤0.05 versus the other three conditions.
As shown in Figure. 4, the wound closure and transwell assays demonstrated that estrogen stimulated the migration/invasion of ABCG2+ cells in a dose-dependent manner. When compared with cells in control culture, the cells treated with 10-9M 17β-estradiol showed a statistically significant increase in the ability of migration and invasion (P<0.05). Furthermore, this effect was greater (P <0.01) in culture with 10-8M 17β-estradiol. However, no statistically significant differences was detected in migration/invasion between cells pretreated with 17 β-estradiol 10-8M and 10-7M (P>0.05). The cells/field was 52.33±4.16 and 57.33±3.06 in the former and latter groups, respectively. Taken together, our results indicate that the high levels of estrogen stimulate the migration of ABCG2+ cells.
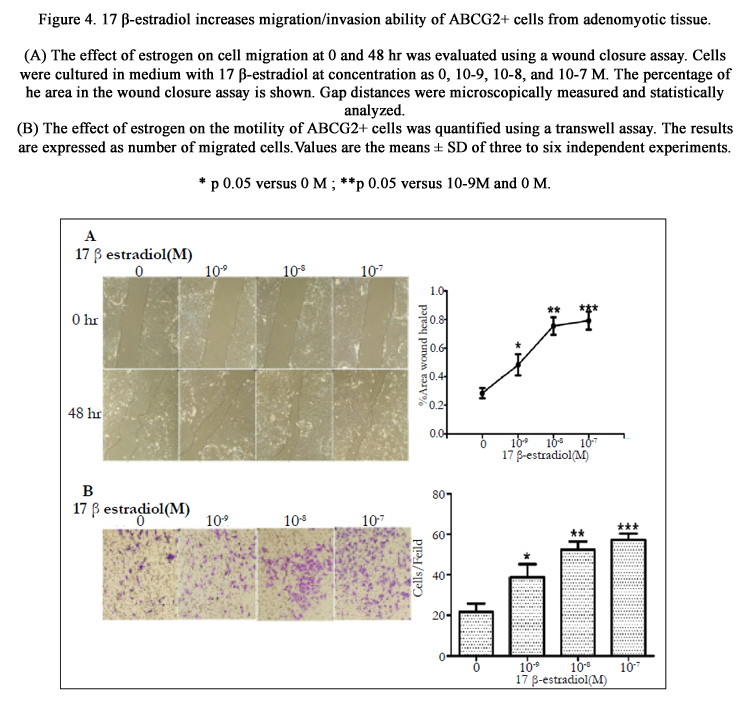
Figure 4. 17 β-estradiol increases migration/invasion ability of ABCG2+ cells from adenomyotic tissue.
(A) The effect of estrogen on cell migration at 0 and 48 hr was evaluated using a wound closure assay. Cells were cultured in medium with 17 β-estradiol at concentration as 0, 10-9, 10-8, and 10-7 M. The percentage of the area in the wound closure assay is shown. Gap distances were microscopically measured and statistically analyzed.
(B) The effect of estrogen on the motility of ABCG2+ cells was quantified using a transwell assay. The results are expressed as number of migrated cells. Values are the means ± SD of three to six independent experiments.
* p<0.05 versus 0 M ; **p<0.05 versus 10-9M and 0 M.
Stem cell hypothesis opens up new avenues for etiology of adenomyosis. Although no special markers are available to distinguish adult stem cells from mature progeny, the presence of endometrium stem cells can be identified by the property of clonogenicity, SP cells and multipotent cells. Herein, we not only showed that human adenomyotic tissues contained SP cells, but also successfully isolated the SP fractions via marker ABCG2+. The clonogenicity and differential potential of ABCG2+ cells are similar to that of endometrial mesenchymal stem-like cells from earlier studies [13, 14]. Our results are consistent with the view that adenomyosis is a disease associated with stem cells [10]. Additionally, the present study shows that estrogen may promote migration/ invasion of ABCG2+ cells.
Endometrium is comprised of surface epithelium and supportive stroma. Some recent studies [15, 16] have demonstrated that two types of stem cells exist in the human endometrium. In the present study, the adenomyotic-derived ABCG2+ cells differentiated into stromal and epithelium cells. We therefore speculate that two distinct types of endometrial epithelial and stromal stem cells may exist in adenomyotic lesion and the stromal stem cells may display more clonogenicity than the epithelium stem cells.
The stem cell niche is a microenvironment of surrounded supporting cells that signal to the stem cells to maintain an undifferentiated state or activate differentiation when necessary [17]. In the present study, ABCG2+ cells display possible stem-cell nature and can differentiate into endometrium-like cells when placed into a favorable conditioned culture. From the different conditions for induced differentiation in the study, we suppose that the most suitable environment is a coexistence of endometrial cells feeder layer and estrogen in culture. The most likely explanation for this finding is that the feeder layer and estrogen provided an optimal stem-cell niche, as it appears that mature endometrial cells assist with the directed differentiation of ABCG2+ cells in an environment with high level of estrogen. Steroid signaling in human is known to be mediated through paracrine mechanisms [18]; therefore, it is reasonable to expect that this action should occur within the specialized environment of a stem cell niche. In the present study, it is likely that the co-cultured ABCG2+ cells and mature endometrial cells in the medium may also produce paracrine factors that are essential for directed differentiation.
Adenomyosis is an estrogen-dependent disease with local production of estrogen, which indicates that estrogen may act indirectly on division-competent cells through paracrine mechanisms. It is possible that ABCG2+ cell fate decisions in adenomyosis may be regulated through steroid hormone action on the niche cells. How can estrogen stimulate differentiation of ABCG2+ cells and their daughter cells? This is clearly an important area deserving future investigation.
Adenomyosis is unique to have benign histology but with malignant characteristics. Epithelial to mesenchymal transition (EMT) allows neoplastic cells to become invasive at the tissue level and subsequently metastasis. Similarly to tumor, development of adenomyosis requires EMT [19]. Local high estrogen concentration in ectopic lesion may be necessary for the maintenance of adenomyosis. The estrogen dependency is often accompanied by the appearance of EMT features, which is a crucial step for the acquisition of invasive properties during adenomyosis progression [20]. Estrogen enhances metastasis and angiogenesis in an adenomyosis model via annexin A2-induced EMT [21]. These data implicate the crucial role of estrogen-induced EMT in the development of adenomyosis. In our study, the transwell experiment is a likely 3D model for stem cell migration/invasion into myometrium. Estrogen exerts a dose-dependent effect on the migration and invasion ability of ABCG2+ cells. The possible explanation for this finding is that estrogen signaling increased nuclear octamer-binding transcription factor 4 (OCT4) expression in stem cells [22]. Previous study has speculated that aberrant expression of Oct-3/4 may contribute to the neoplastic process and play a role in cancer stem cell theory [23]. Study on endometriosis found that the expression of OCT4 was significantly higher in human ectopic endometrial tissues and OCT4 may stimulate the migration activity of endometrial cells [24]. The results of our wound closure and transwell migration assays indicate that estrogen may stimulate ABCG2+ cell invasion and metastasis by EMT process. Thus, our data supports the hypothesis that the increasingly local production of estrogens in adenomyosis changes the stem cell niche microenvironment, leading to enhanced stem cell invasion. Endometrium stem cells reside in the basalis layer flux through the endometrial–myometrial interface and establish endometriotic lesions in the myometrium.
Conclusion
The present study indicates that stem cells are a possible source for adenomyosis. The protein ABCG2+ is a candidate marker for isolating endometrial stem cells and the ABCG2+ cells display stem cell-like properties. Importantly, estrogens play an important role in stem cells migrating and invading into myometrium. Taken together, our data, together with other studies, have enabled us to propose a novel hypothetical pathogenesis model for adenomyosis.
Acknowledgement and Declarations
The study was sponsored by The Science and Technology Committee of Sichuan Province (No. 2012SZ0030) in 2012. The authors declare that they have no competing interests.
References
- Vercellini P, Vigano P, Somigliana E, Daguati R, Abbiati A, et al. (2006) Adenomyosis: epidemiological factors. Best Pract Res Clin Obstet Gynaecol 20(4): 465–77.
- Leyendecker G, Wildt L, Mall G (2009) The pathophysiology of endometriosis and adenomyosis: tissue injury and repair. Arch Gynecol Obstet 280(4): 529 –538.
- Meenakshi M, McCluggage G (2010) Vascular involvement in adenomyosis: report of a large series of a common phenomenon with observations on the pathogenesis of adenomyosis. Int J Gynecol Pathol 29(2): 117–121.
- Huang TS, Chen YJ, Chou TY, Chen CY, Li HY, et al. (2014) Oestrogeninduced angiogenesis promotes adenomyosis by activating the Slug-VEGF axis in endometrial epithelial cells. J Cell Mol Med 18(7): 1358-1371.
- Bulun SE (2009) Endometriosis. N Engl J Med 360: 268– 279.
- Challen GA, Little MH (2006) A side order of stem cells: the SP phenotype. Stem Cells 24(1): 3–12.
- Tsuji S, Yoshimoto M, Takahashi K, Noda Y, Nakahata T, et al. (2008) Side population cells contribute to the genesis of human endometrium. Fertl. Steril 90(4): 1528 –1537.
- Masuda H, Matsuzaki Y, Hiratsu E, Ono M, Nagashima T, et al. (2010) Stem cell-like properties of the endometrial side population: implication in endometrial regeneration. PLoS One 5(4): e10387.
- Chan RW, Ng EH, Yeung ES (2011) Identification of cells with colonyforming activity, self-renewal capacity, and multipotency in ovarian endometriosis. Am J Pathol 178(6): 2832-2844.
- Kato K, Takao T, Kuboyama A, Tanaka Y, Ohgami T, et al. (2010) Endometrial Cancer Side-Population Cells Show Prominent Migration and Have a Potential to Differentiate into the Mesenchymal Cell Lineage. Am J Pathol 176(1): 381-392.
- Chen YJ, Li HY, Chang YL, Yuan CC, Tai LK, et al.(2010) Suppression of migratory/invasive ability and induction of apoptosis in adenomyosisderived mesenchymal stem cells by cyclooxygenase-2 inhibitors. Fertil Steril 94(6): 1972-1979.
- Zhou S, Schuetz JD, Bunting KD, Colapietro AM, Sampath J, et al. (2001) The ABC transporter Bcrp1/ABCG2 is expressed in a wide variety of stem cells and is a molecular determinant of the side-population phenotype. Nat Med 7(9): 1028–34.
- Wu C, Alman BA (2008) Side population cells in human cancers. Cancer Lett 268(1): 1-9.
- Meng X, Ichim TE, Zhong J, Rogers A, Yin Z, et al. (2007) Endometrial regenerative cells: a novel stem cell population. J Transl Med 5(1): 57.
- Schwab KE, Gargett CE (2007) Co-expression of two peri-vascular cell markers isolates mesenchymal stem-like cells from human endometrium. Hum Reprod 22(11): 2903–2911.
- Gargett CE (2007) Uterine stem cells: what is the evidence? Hum Reprod Update 13(1): 87–101.
- Chan RW, Schwab KE, Gargett CE (2004) Clonogenicity of human endometrial epithelial and stromal cells. Biol Reprod 70(6): 1738–1750.
- Moore KA, Lemischka IR (2006) Stem cells and their niches. Science 311(5769): 1880–1885.
- Clarke RB, Howell A, Potten CS, Anderson E (1997) Dissociation between steroid receptor expression and cell proliferation in the human breast. Cancer Res 57(22): 4987–4991.
- Chen YJ, Li HY, Huang CH, Twu NF, Yen MS, et al. (2010) Oestrogeninduced epithelial-mesenchymal transition of endometrial epithelial cells contributes to the development of adenomyosis. J Pathol 222(3): 261–270.
- Ferenczy A (1998) Pathophysiology of adenomyosis. Hum Reprod Update 4(4): 312–322.
- Zhou S, Yi T, Liu R, Bian C, Qi X, et al. (2012) Proteomics identification of annexin A2 as a key mediator in the metastasis and proangiogenesis of endometrial cells in human adenomyosis. Mol Cell Proteomics 11(7): M112.017988.
- Shang Y, Hu X, DiRenzo J, Lazar MA, Brown M (2000) Cofactor dynamics and sufficiency in estrogen receptor-regulated transcription. Cell 103(6): 843–852.
- Tai MH, Chang CC, Olson LK, Trosko JE (2005) Oct4 expression in adult human stem cells: evidence in support of the stem cell theory of carcinogenesis. Carcinogenesis 26(2): 495–502.
- Chang JH, Au HK, Lee WC, Chi CC, Ling TY, et al. (2013) Expression of the pluripotent transcription factor OCT4 promotes cell migration in endometriosis. Fertil Steril 99(5): 1332-1339.





