Effect of Bone Marrow-derived Mesenchymal Stem Cells and Umbilical Cord Blood-CD34+ cells on Experimental Rat liver Fibrosis
Ahmed RH1, Roshdy NK2, Aref MI3, Hassan NM3, Saleh HE1, Mohammed NR1*
1 Zoology Department-Women's College-Ain Shams University, Egypt.
2 Biochemistry Department-Faculty of Medicine-Cairo University, Egypt.
3 Immunology Department-Faculty of Medicine-Al Azhar University, Egypt.
*Corresponding Author
Noura R. Mohammed,
Zoology Department-Women's College-Ain Shams University,
Egypt.
E-mail: noura_rady@yahoo.com
Article Type: Research Article
Received: May 14, 2014; Accepted: May 26, 2014; Published: May 28, 2014
Citation: Mohammed NR, et al. (2014). Effect of Bone Marrow-derived Mesenchymal Stem Cells and Umbilical Cord Blood-CD34+ cells on Experimental Rat liver Fibrosis, Int J Stem Cell Res Transplant, 02(03), 63-68. doi: dx.doi.org/10.19070/2328-3548-1400011
Copyright: Mohammed NR© 2014. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
Background and Objective:Liver disease is one of the major causes of death in many countries. Hence, the development of effective therapies for liver fibrosis is a major aim of medical research. So this study was designed to investigate the therapeutical role of mesenchymal stem cells (MSCs) and hematopoietic stem cells (HSCs) transplantation in the experimental rat liver fibrosis.
Design and Method: Bone marrow-derived MSCs were isolated from femoral and tibial bones of male albino rats, then were grown and propagated in culture for 2 weeks and were characterized morphologically and by detection of CD29 by real time-PCR. Human umbilical cord blood cells were obtained after full-term caesarean delivery from healthy donors after written informed consent. Low-density mononuclear cells were separated over Ficoll- Paque (Gibco-Invitrogen, Grand Island, NY), and then CD34+ HSC was isolated using a magnetic cell sorter (MiniMACS; Miltenyi Biotec, Bergisch Gladbach, Germany). The cells were then infused intraperitoneally in rats that received CCl4 injection to induce liver fibrosis. Rats were divided into 4 groups: control, CCl4, CCl4 plus MSC, and CCl4 plus CD34+. Liver tissue was examined histopathologically for all groups. The expression of collagen I and metalloproteinase-2 genes as a marker of liver fibrosis was measured by real time RT- PCR.
Results: The results of the present study showed that both MSCs and CD34+ have a significant antifibrotic effect as evidenced by the significant decrease in liver collagen gene expression as well as the decrease in MMP-2 (p < 0.05) compared to the CCl4 group.
2.Introduction
3.Materials and Methods
3.1.Preparation of BM-derived MSCs
3.2.Isolation of CD34+ stem cells
3.3.Labeling of stem cells with PKH26
3.4.CCl4-induced liver fibrosis model and MSC administration
3.5.Analysis of liver histopathology
3.6.Quantitave RT-PCR analysis for collagen I and MMP-2
4.Results
4.1.Liver histopathology
4.2.The collagen I gene expression
4.3.The MMP-2 gene expression
4.4.Correlation between type 1 collagen and MMP-2 gene expression
5.Discusion
6.Conclusion
7.References
Keywords
Liver Fibrosis; Hematopoietic Stem Cells; Mesenchymal Stem Cells.
Introduction
Hepatic fibrosis (HF) is the accumulation of extracellular matrix, or scar, in response to acute or chronic liver injury. Fibrogenesis represents a wound healing response to injury, and ultimately leads to cirrhosis. Cirrhosis is the end-stage consequence of fibrosis of the hepatic parenchyma, resulting in nodule formation that may lead to altered hepatic function and blood flow [1]. Alcohol abuse, chronic viral hepatitis (HBV and HCV), obesity, autoimmune hepatitis, parasitic diseases (i.e. schistosomiasis), metabolic disorders (hemochromatosis and wilson's disease), biliary disease, persistent exposure to toxins and chemicals, and drug-induced chronic liver diseases are the most common causes of hepatic fibrosis [2]. Hepatic stellate cells are the major source of fibrillar collagens and other ECM proteins that characterize liver fibrosis [3]. As the liver becomes fibrotic, the total content of collagens and non-collagenous components increases three-to fivefold, accompanied by a shift in the type of ECM in the subendothelial space from the normal low-denisty basement membrane-like matrix to interstitial-type matrix [4]. This 'capillarization' leads to the loss of hepatocyte microvilli and the disappearance of endothelial fenestration. MMPs are the main enzymes responsible for ECM degradation and TIMPs have the ability to inhibit MMPs [5]. Mesenchymal stem cells (MSCs) have been isolated from several sources, BM, cord blood, amniotic fluid, placenta and adipose tissue, and therefore, become a prime candidate for cell therapy. Several studies have demonstrated that MSCs isolated and expanded in vitro can be induced to differentiate into hepatocyte-like cells [6]. HSCs obtained from cord blood have been shown to be a suitable alternative to adult bone marrow or peripheral blood in transplants for the treatment of leukemia, lymphoma, aplastic anemia, and inherited disorders of immunity and metabolism [7]. Traditionally HSCs generate all mature blood cell types of the hematolymphatic system. More recently, HSCs have shown to display remarkable plasticity and can apparently differentiate into several non-hemolymphatic tissue lineages [8]. Therefore, UBCs may become a hopeful candidate for a hepatocyte progenitor source like BMCs. Therefore, the aim of the present work was to investigate the therapeutical role of stem cell transplantation in the experimental rat liver fibrosis and compare the proliferative potency of them.
Materials and Methods
Bone marrow was harvested by flushing the tibiae and femurs of 6 weeks old male white albino rats, weighing between 140 and 160g, with Dulbecco’s modified Eagle’s medium (DMEM, GIBCO/BRL) supplemented with 10% fetal bovine serum (GIBCO/ BRL). Nucleated cells were isolated with a density gradient [Ficoll/Histopaque (Pharmacia)] and resuspended in complete culture medium supplemented with 1% penicillin-streptomycin (GIBCO/BRL). Cells were incubated at 37°C in 5% humidified CO2 for 12-14 days. Media was changed every 3-4 days. When large colonies developed (80-90% confluence), cultures were washed twice with phosphate buffer saline (PBS) and cells were trypsinized with 0.25% trypsin in 1mM Ethylene Diamine Tetra Acetate (EDTA) (GIBCO/BRL) for 5 minutes at 37°C. After centrifugation, cell pellets were resuspended with serum-supplemented medium and incubated in 50 cm2 culture flask (Falcon). The resulting cultures were referred to as first-passage cultures [9]. MSCs in culture were characterized by their adhesiveness and fusiform shape [10]. We also detected CD29 gene expression by RT-PCR as a marker of MSCs [11].
Human Umbilical Cord Blood (UCB) was used for separation of mononuclear cells (MNCs) after obtaining an informed consent and research ethics committee approval. Anticoagulated cord blood was diluted 1:4 with PBS containing 2 mM EDTA (Gibco-Invitrogen, Grand Island, NY) and 35 ml of the diluted sample was carefully layered on 15 ml Ficoll-Paque (Gibco-Invitrogen, Grand Island, NY), then they were centrifuged for 35 min. at 400xg rpm. CD34+ HSC were isolated from UCB using MACS (magnetic cell sorting) kits (MiniMACS; Miltenyi Biotec, Bergisch Gladbach, Germany) according to Gordon et al. 2006 [12].
Two types of stem cells were labeled with PKH26, which is a red fluorochrome. It has excitation (551nm) and emission (567nm) characteristics compatible with rhodamine or phycoerythrin detection systems. The linkers are physiologically stable and show little to no toxic side-effects on cell systems. Labeled cells retain both biological and proliferating activity, and are ideal for in vitro cell labeling, in vitro proliferation studies and long, in vivo cell tracking. In the current work, MSCs were labeled with PKH26 from Sigma Company (Saint Louis, Missouri USA). Cells were centrifuged and washed twice in serum free medium. Cells were pelleted and suspended in dye solution. Cells were injected intraperitoneally in rats. After one month, liver tissue was examined with a fluorescence microscope to detect and trace the cells.
Thirty male white Albino rats were 6 weeks old, weighing between 150 and 200 g constituted the materials of this work. The animals were housed in well ventilated hygienic experimental animal house under constant environmental and nutritional conditions. Liver fibrosis was induced by CCl4 injected by subcutaneous route at a dose of 0.2 mL/100 g body weight of 40 mL/L CCl4 (99 % purity,Sigma, St Louis, USA) dissolved in equal volume of castor oil (Sigma, St. Louis, USA). The injection was given twice a week for 6 weeks according to Zhao et al., 2005 [13]. The same volume of castor oil alone was used as a control. The delay in administration of stem cells until 6 weeks of injection of CCl4 was suggested by histopathological examination of liver samples and also supported by the work of Zhao et al (2005). Stem cells were given at a dose of 107 cells per rat to establish humanized rat xenograft model. On day 0, rats were divided into the following groups:
1. Control group: 10 rats received 0.2 mL/100 g body weight of castor oil twice a week for 6 weeks.
2. CCl4 group: 10 rats received 0.2 mL/100 g body weight of CCl4 by the schedule mentioned above. Liver fibrosis was determined by histopathological examination.
3. CCl4/MSCs group: involved 10 rats that received 0.2 ml/100 g body weight of CCl4 by the schedule mentioned above. The rats were infused with a dose of 107 undifferentiated cells per rat intraperitoneally (IP) according to Zhao et al [14].
4. CCl4/CD34+ group: involved 10 rats that received 0.2ml/100 g body weight of CCl4 by the schedule mentioned above. The rats were infused with a dose of 107 undifferentiated cells per rat intraperitoneally (IP) according to Di Campli et al [15].
Liver samples were collected into PBS and fixed overnight in 40g/L paraformaldehyde in PBS at 4°C. Serial 5-μm sections of the right lobes of the livers were stained with hematoxylin and eosin (HE) and were examined histopathologically.
Total RNA was extracted from liver tissue homogenate using RNeasy purification reagent (Qiagen, Valencia, CA). cDNA was generated from 5 μg of total RNA extracted with 1 μl (20 pmol) antisense primer and 0.8 μl superscript AMV reverse transcriptase for 60 min at 37°C. The relative abundance of mRNA species was assessed using the SYBR® Green method on an ABI prism 7700 sequence detector system (Applied Biosystems, Foster City, CA) [16]. PCR primers were designed according to Morimoto et al. 2011 [17]. All primer sets had a calculated annealing temperature of 60°. Quantitative RT-PCR was performed in duplicate in a 25-μl reaction volume consisting of 2X SYBR Green PCR Master Mix (Applied Biosystems), 900 nM of each primer and 2–3 μl of cDNA. Amplification conditions were 2 min at 50°, 10 min at 95°and 40 cycles of denaturation for 15 s and annealing/extension at 60° for 10 min. Data from real-time assays were calculated using the v1•7 Sequence Detection Software from PE Biosystems (Foster City, CA). Sequence of the murine primers used for realtime PCR:
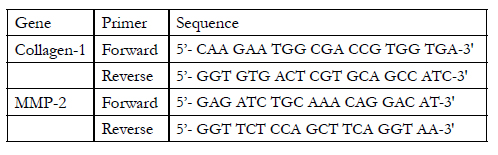

Table
Statistical analysis:Data are expressed as Mean ± SD. Significant differences were determined using ANOVA and post hoc tests for multiple comparisons using SPSS 15.0 computer Software. P values of <0.05 were considered significant.
Results
Histopathological examination of liver tissue showed that two types of stem cells have a significant antifibrotic effect as evidenced by the decrease in liver extracellular matrix (ECM) compared to the CCl4 group (Fig. 1).
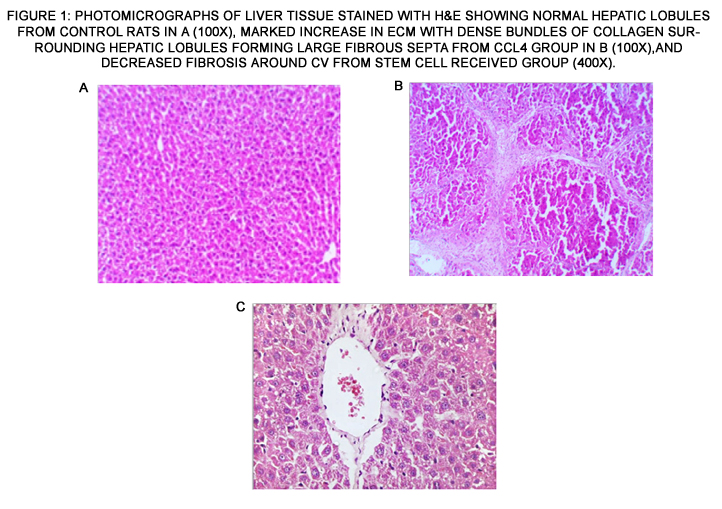
Figure 1: Photomicrographs of liver tissue stained with H&E showing normal hepatic lobules from control rats in a (100x), marked increase in ECM with dense bundles of collagen surrounding hepatic lobules forming large fibrous septa from CCl4 group in b (100x),and decreased fibrosis around CV from stem cell received group (400x).
There was a significant difference (p<0.05) in gene expression of all groups compared to normal group. CCl4, MSCs,and CD34+ groups showed a significant increase (1.02±0.26,0.50±0.21 & 0.51±0.23) respectively, compared to the normal group (0.12±0.03). Meanwhile, stem cell received groups showed a significant decrease in gene expression compared to the CCl4 group. Also, there was no significant difference between stem cell received groups (Fig.2).
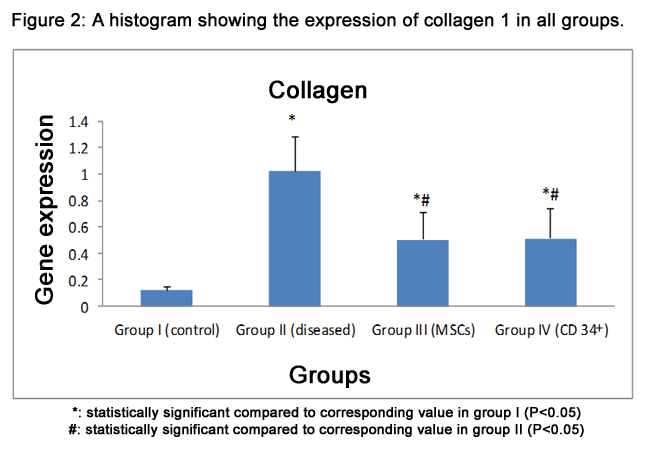
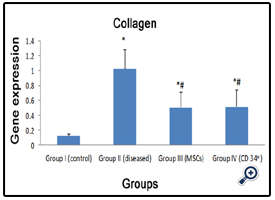
Figure 2: A histogram showing the expression of collagen 1 in all groups.
*: statistically significant compared to corresponding value in group I (P<0.05)
#: statistically significant compared to corresponding value in group II (P<0.05)
MMP-2 mRNA was significantly increased (p<0.05) in fibrotic liver group and CD34+ treated group (0.91±0.05 & 0.46±0.18)compared to normal group (0.24±0.06). On the other hand, MMP-2 gene expression was significantly decreased (p<0.05) in the MSCs group (0.40±0.18) and CD34+ group compared to CCl4 group, and there was no significant difference between stem cell received groups (Fig.3).
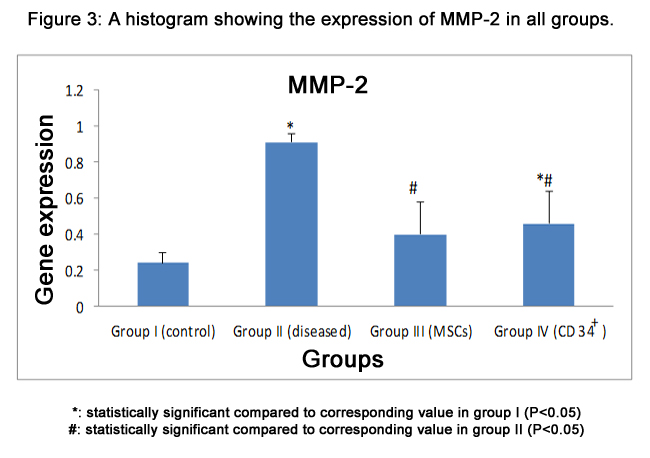
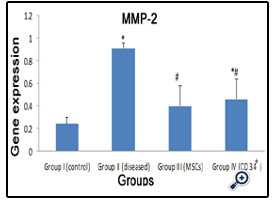
Figure 3: A histogram showing the expression of MMP-2 in all groups.
*: statistically significant compared to corresponding value in group I (P<0.05)
#: statistically significant compared to corresponding value in group II (P<0.05)
Using pearson correlation, the results showed that a strong positive correlation (p<0.05) in the expression of both genes. When the level of collagen 1 expression increased, the level of MMP-2 expression also increased (R. linear) as illustrated in figure (4).
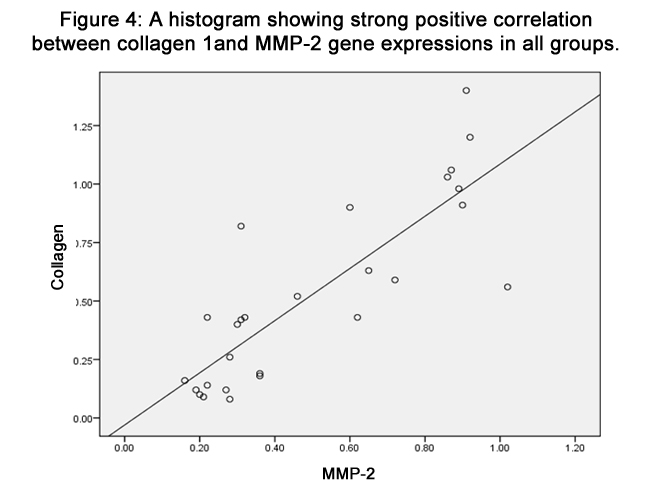
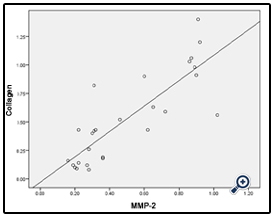
Figure 4: A histogram showing strong positive correlation between collagen 1and MMP-2 gene expressions in all groups.
Discusion
The liver is responsible for many critical functions in the body and loss of those functions may leads to damage of the body. Hepatic fibrosis (HF) represents a scarring response to either acute or chronic cellular injury. The scar is typically composed of fibrillar collagen type I and III, proteoglycans, fibronectin and hyaluronic acid [18]. As a result, alteration in the physiological architecture of the liver occurs, particularly in the space of Disse, where the low electron-dense ECM is replaced by one rich in fibrillar collagens and fibronectin. This leads to loss of endothelial cell fenestrations, impaired exchange of solutes among neighboring cells, altered hepatocyte function and subsequent parenchymal cell damage [19]. Carbon tetrachloride-induced fibrosis and cirrhosis is one of the oldest and probably the most widely used toxinbased experimental model for the induction of fibrosis. It has the advantages that it has been clearly characterized and in many respects mirrors the pattern of disease seen in human fibrosis and cirrhosis associated with toxic damage [20,21]. In this study, when rats were treated with CCl4 twice weekly for 6 weeks, the liver exhibited a marked increase in extracellular matrix (ECM) content and displayed bundles of collagen surrounding the lobules leading to large fibrous septa. All of these are characteristics of advanced fibrosis. Meanwhile, histopathologic examination of livers showed decreased fibrosis after administration of MSCs and CD34+ stem cells, this in agreement with Abdel Aziz et al. [22] and Di Campli et al. [15]. Also the molecular results in this study, detected a significant increase in gene expression of collagen type I and MMP-2mRNA in rats given CCl4 than rats with only castor oil, and this with agreement with the study of Abdel Aziz et al. [22] and Kollet et al. [23]. Liver fibrosis refers to the accumulation of extracellular matrix (ECM) proteins, which occurs in most types of chronic diseases. Of these proteins, collagen, which is the body’s self-repairing protein induced by inflammation, most commonly accumulates [24]. At the early stage of liver fibrosis, collagen type IV increases first and then gradually decreases, while the collagen type III increases and expresses predominantly. In the developing and advanced stages, the collagen type I gradually increased, and collagen of both type I and II significantly increased, especially the type I [25,26,27]. In the progression of liver fibrosis, MMP-2 is also involved in the degradation of fibrotic matrix [28]. An interesting recent observation is that HSCs-mediated activation of MMP-2 is significantly induced in the presence of collagen type I (the principal matrix protein found in fibrotic liver) [29]. This could contribute to further degradation of the normal liver matrix, leading to increased activation of HSCs and increased synthesis of type I collagen. This positive feedback loop would theoretically promote progression of liver fibrosis [30]. Stem cell-based cytotherapy has shown promising benefit on animal models and some clinical patients [31]. The result of the present work showed strong red auto fluorescence after transplantation in rats, confirming that these cells were actually insinuating themselves into the liver tissue as detected by fluorescent microscope. Molecular mechanisms underlying stem cells mobilization and homing into the injured liver are still poorly understood [32]. Naturally, the migration, differentiation and growth are mediated by the tissue, degree of injury and SCs involved. Damaged tissue releases factors that induce SCs homing. In our study, quantitative RT-PCR analysis detected a significant reduction in the indicators of liver fibrosis such as collagen I and MMP-2 gene expression showed decrease in mRNA level after MSCs administration and this in agreement with Abdel Aziz et al. [33]. In an attempt to reveal the underlying mechanism, Prosser et al. demonstrated that
(a) MSCs were able to produce hepatocyte growth factor (HGF), which in turn could promote the MSCs to undergo the process of trans-differentiation into parenchymal hepatocyte [34].
(b) MSCs prevent the liver from undergoing fibrogenesis via secreting a variety of cytokines, such as HGF, interleukin (IL)-6 and -10 and so on. These cellular factors may inhibit hepatic fibrosis through multiple mechanisms.
(c) MSCs may dissolve fibrosis directly. There was evidence suggesting that MSCs were able to produce the matrix metalloproteinase (MMPs), an enzyme capable of degrading the extracellular matrix, which alleviates hepatic cirrhosis directly [35]. On the other hand, umbilical cord blood contains a higher amount of hematopoietic stem cells compared with bone marrow. The immaturity of umbilical cord blood stem cells (UCBSCs) affords them numerous advantages over adult bone marrow-derived stem cells (BMSCs) in cell transplantation [36]. Controversy has recently arisen as to whether HSCs contribute to the hepatocyte lineage in liver injury via transdifferentiation alone or by adopting the phenotype of hepatocytes after spontaneous cell fusion [37]. Recent reports in favor of the fusion hypothesis have demonstrated that adult cells can adopt the phenotype of other cell lines by fusing with embryonal stem cells [38,39] as well as BM-derived hepatocytes generated by in vivo cell fusion [40]. In support of transdifferentiation, several groups have demonstrated that HSCs can differentiate into hepatocytes [8,41] nd pancreatic endocrine cells [42] without any evidence of cell fusion. Whatever the mechanism of hepatic regeneration is, the trafficking of HSCs to the liver may play an important component of the reparative process in liver injury [43]. Finally, where there are controversial about the mechanism by which MSCs repair liver fibrosis, our study tend to that BM-derived stem cells exert a paracrine proliferative effect on endogenous hepatocytes, as evidenced by studies of experimental animals and humans. The self-renewal potential of endogenous hepatocytes in cirrhotic liver is compromised, and they might acquire replicative senescence after many rounds of injury and repair [44,45].
Conclusion
Transplantation of BM-MSCs and UCB-HSCs ameliorate CCl4-induced liver fibrosis in rats mediated by the improvement of fibrogenesis by decreasing expression of collagen type I and subsequently MMP-2 gene expression. Furthermore, studies with longer follow up are required to confirm the safety of this therapy.
References
- Ahmad A, Ahmad R(2012) Understanding the mechanism of hepatic fibrosis and potential therapeutic approaches. Saudi J Gastroenterol.. 18: 155 67.
- Mormone E, George J and Nieto N (2011) Molecular pathogenesis of hepatic fibrosis and current therapeutic approaches. Chemico-Biological Interactions; 193: 225-231.
- Friedman S.L(2008) Mechanisms of hepatic fibrogenesis. Gastroenterol.; 134: 1655-1669.
- Schuppan D, Ruehl M, Somasundaram R. and Hahn E.G (2001) Matrix as modulator of stellate cell and hepatic fibrogenesis. Semin. Liver Dis; 21: 351-372.
- Goto T, Mikami K.I, Miura K, Ohshima S, Yoneyama K, Nakane K, Watanabe D, Otaka M. and Watanabe S (2004) Mechanical stretch induces matrix metalloproteinase 1 production in human hepatic stellate cells. Pathophysiology; 11(3): 153-158.
- Banas A, Teratani T, Yamamoto Y, Tokuhara M, Takeshita F, Osaki M, Kato, Okochi H and Ochiya T(2009) Rapid hepatic fate specification of adiposederived stem cells and their therapeutic potential for liver failure. J. Gastroenterol. Hepatol.; 24: 70-77.
- Talens R, Bonora A and Jover R( 2006) Hepatogenic differentiation of human mesenchymal stem cells from adipose tissue in comparison with bone marrow mesenchymal stem cells. World J. Gastroenterol.; 12: 5834-5845.
- Newsome P.N, Johannessen I, Boyle S, Dalakas E. and Plevris J.N(2003) Human cord blood-derived cells can differentiate into hepatocytes in the mouse liver with no evidence of cellular fusion. Gastroenterology; 124(7): 1891-1900.
- Alhadlaq A and Mao J.J(2004) Mesenchymal stem cells: isolation and therapeutics. Stem Cells Dev. J; 13: 436-448.
- Rochefort G.Y, Vaudin P, Bonnet N, Pages J.C, Domenech J and Charbord P(2005)Influence of hypoxia on the domiciliation of mesenchymal stem cells after infusion into rats: possibilities of targeting pulmonary artery remodeling via cells therapies. Respir. Res; 6: 125.
- Munoz-Fernandez R, Blanco F.J, Frecha C, Martin F, Kimatrai M(2006) Follicular dendritic cells are related to bone marrow stromal cell progenitors and to myofibroblasts. J. Immunol; 177: 280-289.
- Gordon M.Y, Levicar N, Pai M, Bachellier P, Dimarakis I(2006) Characterization and clinical application of human CD34+ stem/progenitor cell population mobilized into the blood by granulocyte colony-stimulating factor. Stem Cells; 24: 1822-1830.
- Zhao D.C, Lei J.X, Chen R, Yu W.H, Zhang X.M(2005)Bone marrowderived mesenchymal stem cells protect against experimental liver fibrosis in rats. World J Gastroenterol; 11(22): 3431-3440.
- Zhao W, Li J.J, Cao D.Y, Li X, Zhang L.Y(2012)Intravenous injection of mesenchymal stem cells is effective in treating liver fibrosis. World J. Gastroenterol; 18(10): 1048-1058.
- Di Campli C, Piscaglia A.C, Pierelli L, Rutella S, Bonanno G(2004) A human umbilical cord stem cell rescue therapy in a murine model of toxic liver injury. Digestive and Liver Disease; 36: 603-613.
- Singh K.P, Gerard H.C, Hudson A.P. and Boros D.L(2004) Expression of matrix metalloproteinases and their inhibitors during the resorption of schistosome egg-induced fibrosis in praziquantel-treated mice. Immunology; 111(3): 343-352.
- Morimoto Y, Oyabu T, Ogami A, Myojo T, Lenggoro W(2011) Investigation of gene expression of MMP2 and TIMP-2 mRNA in rat lung in inhaled nickel oxide and titanium dioxide nanoparticles. Industrial Health; 49: 344- 352.
- George J, Tsutsumi M and Takase S(2004) Expression of hyaluronic acid in N-nitrosodimethylamine induced hepatic fibrosis in rat. Int. J. BioChem. Cell Biol.; 36(2): 307-319.
- Hernandez-Gea V. and Freiedman S.L. (2011): “Pathogenesis of liver fibrosis”. Annu. Rev. Pathol. 6: 425-456.
- Tsukamoto H., Matsuoka M. and French S.W(1990) Experimental models of hepatic fibrosis. Semin. Liver Dis; 10(1): 56-65.
- Perez Tamayo R( 1983) Is cirrhosis of the liver experimentally produced by CCl4 and adequate model of human cirrhosis. Hepatology; 3(1): 112-20.
- Abdel Aziz M.T, Atta H.M, Mahfouz S, Fouad H.H, Roshdy N.K (2007) Therapeutic potential of bone marrow-derived mesenchymal stem cells on experimental liver fibrosis. Clinical Biochemistry Med; 40: 893-899.
- Kollet O, Shivtiel S, Chen Y.Q, Suriawinata J, Spiegel A(2003) HGF, SDF-1, and MMP-9 are involved in stress-induced human CD34+ stem cell recruitment to the liver. J. Clin. Invest; 112: 160-169.
- Li W, Ren G, Huang Y, Su J, Han Y(2012)Mesenchymal stem cells: a double-edged sword in regulating immune responses. Cell Death Differ; 19(9): 1505-13.
- Du W.D, Zhang Y.E, Zhai W.R and Zhou X.M( 1999) Dynamic changes of type I, II and IV collagen synthesis and distribution of collagen producing cells in carbon tetrachloride induced rat liver fibrosis. World J. Gastroentero; 5: 397-403.
- Nakatsukasa H, Evarts R.P, Hsia C.C. and Thorgeirsson S.S(1990) Transforming growth factor-β1 and type I procollagen transcripts during regeneration and early fibrosis of rat liver. Lab. Invest; 63: 171-179.
- Wang Y.J, Sun Z.Q, Quan Q.Z, Zhang Z.J, Liu X.F(1994) Study on type IV collagen and fat storing cells in fibrotic rat liver treated by tetrandrine. Xin Xiaohuabingxue Zazhi; 2: 78-79.
- Kerkvliet E.H, Docherty A.J, Beertsen W and Everts V(1999) Collagen breackdown in soft connective tissue explants is associated with the level of active gelatinase A (MMP-2) but not with collagenase. Matrix Biol.; 18: 373-380.
- Theret N, Lehti K, Musso O and Clement B(1999) MMP2 activation by collagen I and concanavalin A in cultured human hepatic stellate cells. Hepatology; 30: 462-468.
- Arthur M.J(2000) Fibrogenesis II metalloproteinases and their inhibitors in liver fibrosis. Am. J. Physiol. Gastrointest. Liver Physiol.; 279: G245-G249.
- Dai L.J, Li H.Y, Guan L.X, Ritchie G. and Zhou J.X(2009) The therapeutic potential of bone marrow-derived mesenchymal stem cells on hepatic cirrhosis. Stem Cell Research; 2: 16-25.
- Muraca M(2011) Evolving concepts in cell therapy of liver disease and current clinical perspectives. Digestive and Liver Disease; 43(3): 180-187.
- Abdel Aziz M.T., Atta H.M., Roshdy N.K., Rashed L.A., Sabry D., Hassouna A.A., Aboul Fotouh G.I., Hasan N.M., Younis R.H. and Chowdhury J.R. Amelioration of murine schistosoma mansoni induced liver fibrosis by mesenchymal stem cells. Journal of Stem Cells and Regenerative Medicine 2012; 8(1): 28-34.
- Prosser C.C, Yen R.D and Wu J(2006)Molecular therapy for hepatic injury and fibrosis; where are we. World J. Gastroenterol; 12(4): 509-15.
- Hardjo M, Miyazaki M, Sakaguchi M, Masaka T, Ibrahim S(2009) Suppression of carbon tetrachloride-induced liver fibrosis by transplantation of a clonal mesenchymal stem cell line derived from rat bone marrow. Cell Transplant; 18(1): 89-99.
- Mayani H and Lansdorp P.M(2009) Biology of human umbilical cord bloodderived hematopoietic stem/progenitor cells. Stem Cells 1998; 16: 153-165.
- Austin T.W and Lagasse E(2003) Hepatic regeneration from hematopoietic stem cells. Mechanisms of Development; 120(1): 131-135.
- Terada N, Hamazaki T, Oka M, Hoki M, Mastalerz D.M(2002)Bone marrow cells adopt the phenotype of other cells by spontaneous cell fusion. Nature; 416: 542-545.
- Ying Q.L, Nichols J, Evans E.P and Smith A.G(2002)Changing potency by spontaneous fusion. Nature; 416: 545-548.
- Wang X, Willenbring H, Akkari Y, Torimaru Y, Foster M(2003) Cell fusion is the principal source of bone marrow-derived hepatocytes. Nature; 422: 897-901.
- Ishikawa F, Drake C.J, Yang S, Fleming P, Visconti R.P(2003)Transplanted human cord blood cells give rise to hepatocytes in engrafted mice. Acad. Sci.; 996: 174-185.
- Ianus A, Holz G.G, Theise N.D and Hussain M.A(2003) In vivo derivation of glucose-competent pancreatic endocrine cells from bone marrow without evidence of cell fusion. J. Clin. Invest; 111: 843-850.
- Dalakas E, Newsome P.N and Harrison D.J(2005) Hematopoietic stem cell trafficking in liver injury. FASEB J; 19: 1225-31.
- Fassett J.T, Tolbot D, Nelsen C.J, Albrecht J.H and Hansen L.K(2003) The role of collagen structure in mitogen stimulation of ERK, cyclin D1 expression, and G1-S progression in rat hepatocytes. J. Biol. Chem; 278: 31691- 1700.
- Trak-Smayra V, Contreras J, Dondero F, Durand F, Dobois S(2004) Role of replicative senescene in the progression of fibrosis in hepatitis C virus (HCV) recurrence after liver transplantation.Transplantation 77: 1755-1760.





