Active Hexose Correlated Compound and T Cell Response in Hind - Limb - Unloaded BALB/c Mice
Kogiso M1,2, Wakame K1,3, Sakai T2, Yamamoto S5, Sundaresan A4, Kulkarni AD1*
1 Department of Surgery, The University of Texas Health Science Center and GSBS at Houston, Houston, USA.
2 International Public Health Nutrition, Institution of Health Biosciences, The University of Tokushima, Tokushima, Japan.
3 Amino Up Chemical Co., (current Hokkaido Pharmaceutical University) Sapporo, Japan.
4 Department of Biology, Texas Southern University, Houston, Texas.
5 Graduate School of Human Life Sciences, Jumanji University, Japan.
*Corresponding Author
Anil D. Kulkarni MSc, PhD,
Department of Surgery, The University of Texas Health Science Center and GSBS at Houston,
Houston, 77030, USA.
Tel: 713-500-7303
Fax: 713-500-7268
E-mail: anil.d.kulkarni@uth.tmc.edu
Received: November 05, 2015; Accepted: December 09, 2015; Published: December 14, 2015
Citation: Kulkarni AD et al., (2015) Active Hexose Correlated Compound and T Cell Response in Hind - Limb - Unloaded BALB/c Mice. Int J Surg Res 2(5) 32-38. doi: dx.doi.org/10.19070/2379-156X-150007
Copyright: Kulkarni AD© 2015. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
Introduction: Microgravity - induced immune dysfunction is a potential risk for infection and cancer in astronauts. Active hexose correlated compound (AHCC), a compound obtained from fermented extract of the mycelia of Basidiomycetes mushrooms, has immunomodulating effects. As a nutritional countermeasure, we examined the effect of AHCC on microgravity- induced immune changes by using a hind-limb unloading (HLU) of mice as a microgravity analog.
Methods: BALB/c mice were divided into non-HLU and HLU groups. Each group was further divided into subgroups receiving drinking water containing 0, 0.1, 1, and 10 g/kg/day AHCC. After 7 days of HLU and feeding treatment, the proliferative response of popliteal lymph nodes (PLN) to alloantigen was assessed to evaluate in vivo T cell function. Furthermore, LPS-stimulated inflammatory cytokine and chemokine production in splenocytes was examined.
Results: The response of PLN was significantly decreased in the HLU group and AHCC supplementation significantly reversed the response. LPS-stimulated cytokine and chemokine production was increased in the HLU group compared to that in the non- HLU group, and AHCC supplementation tended to increase those levels further.
Discussion: AHCC enhances in vivo T cell function in microgravity, suggesting that AHCC supplementation under microgravity may have a beneficial role in maintenance of the immune system.
2.Introduction
3.Materials and Methods
3.1 Animals
3.2 Administration of AHCC
3.3 HLU procedure
3.4 Popliteal lymph node (PLN) assay (in vivo proliferation)
3.5 Splenocyte proliferation assay (in vitro proliferation)
3.6 Splenocyte cytokine production
3.7 Statistical analysis
4.Results
4.1 Effect of HLU and AHCC supplementation on in vivo host immune response
4.2 Effect of HLU and AHCC supplementation on in vitro T cell function
4.3 Effect of HLU and AHCC supplementation on cytokine and chemokine production
5.Discussion
6.Acknowledgments
7.References
Keywords
AHCC; Hind - Limb Unloading; T Cell Function; Inflammatory Response.
Introduction
Space travelers have been exposed to numerous psychological and physical stressors. Physical stressors include microgravity, radiation, malnutrition, and microbial contamination and atmospheric pollutants due to recycled air and water [1, 2]. Microgravity is one of the most detrimental stressors for space travelers and aggravates bone structure and composition, immune function and the psychoneuroendocrine system and induces muscle atrophy.
Spaceflight causes various changes in the immune system, including decreases in T cell proliferation, cell-mediated activity, natural killer (NK) cell activity, macrophage function and responses of bone marrow cells to colony stimulating factors and alteration in cytokine production [1-3]. However, because of the limited sample number, different mission durations and different experimental conditions, spaceflight-induced immune changes have not been fully elucidated and ground-based studies have also been required.
Hind-limb unloading (HLU) of rodents is widely used as a ground-based model for modeling some effects of microgravity [4]. Some of the physiological changes induced by HLU parallel those induced by spaceflight [5]. These physiological changes include change in bone structure and composition, muscle atrophy, fluid shift to the head, and changes in the immune system. Studies on changes in the immune system of rodents subjected to HLU have revealed decreased T cell proliferation, decreased cellmediated activity, decreased ability to kill phagocytosed bacteria, decreased responses of bone marrow cells to colony stimulating factors, and altered cytokine production [3, 6-8]. Immune dysfunction results in a state of vulnerability to infection and increased cancer risk [9]. Microgravity-induced immune dysfunction results in increased risks and susceptibility to infection and cancer. In rodents subjected to HLU, decreased resistance to infection [9-11] and increased tumor growth [12] have been reported. Since the immune system closely functions with nervous, endocrine and other systems, immune dysfunction may cause many problems systemically for humans. There are no known countermeasures for immune dysfunction in microgravity environment to prevent infectious complications. Besides, the efficacy of antibiotics is known to be reduced and microbial mutation rates are dramatically increased in microgravity [13]. Furthermore, antibiotics may have adverse effects. The possibility of improvement in immune dysfunction by nutritional supplementation is an issue of interest because there should be no adverse effects and a nutritional countermeasure is cost-effective compared to other therapies [14, 15]. The levels of some nutrients that are synthesized in animals and humans are insufficient under stress conditions, including microgravity. Kulkarni et al and Yamauchi et al have examined the effect of nucleotide supplementation in rodents subjected to HLU and have reported beneficial effects on immune function [8, 16]. However, there is scanty-to-nil literature for immunonutritional countermeasure and the effect of non-inherent nutritional substrate supplementation has not been studied well.
We examined the effect of a novel nutritional substrate active hexose correlated compound (AHCC) on immune function in microgravity analog. AHCC is a compound obtained from enzymefermented extract of the mycelia of Basidiomycetes mushrooms. Chemical analysis has revealed that AHCC consists of oligosaccharides, amino acids, lipids, and minerals [17]. The main components of AHCC are oligosaccharides (~74% of AHCC), and approximately 20% of AHCC are partially acetylated α-1,4-glucans with a mean molecular weight of 5 kDa. These oligosaccharides including α-1,4-glucans are believed to be the active components of AHCC [18, 19].
AHCC has beneficial immunomodulating effects in both innate and acquired immunity in animals and humans. Enhanced NK activity [19] and modulation of inflammatory and T helper (Th) 1/Th2 cytokine production [7, 18] and antibody production [10] have been reported. These effects result in a good clinical outcome. Increased resistance to infection [10, 19, 20], anti-tumor effects [21], anti-inflammatory effect in colitis [22], and improvement of quality of life in cancer patients [23] have been reported without adverse effects. These findings suggest that AHCC supplementation may have a beneficial effect on microgravityinduced immune dysfunction. It has been reported that AHCC improves resistance to infection [10] and modulates inflammatory cytokine production [7] in HLU. It is also reported that in vitro Th1 cytokine production in splenocytes from AHCC-fed HLU mice was increased [7]. However, the effect of AHCC supplementation on microgravity-induced immune dysfunction, especially in vivo T cell dysfunction, has not been fully elucidated. In this study, we examined the effect of AHCC supplementation on microgravity-induced immune dysfunction and found that AHCC supplementation has a beneficial role in maintenance of the immune system, especially in vivo T cell function.
Materials and Methods
The Animal Care and Facilities Use Committee at The University of Texas approved the animal protocol. The experiments were performed in adherence to the National Institutes of Health Guidelines on the Use of Laboratory Animals. Seven - to eight week-old female BALB/c mice and C57BL/6 mice were obtained from Harlan Sprague Dawley, Inc. (Indianapolis, IN). Mice were maintained under a 12-h light:dark cycle at 25 ± 2°C and 50 to 70% relative humidity. The mice were allowed to adapt to our laboratory environment for 1 week before the start of the experiment, during which time they were provided with a commercial rodent diet and water ad libitum.
After the period of acclimatization, the BALB/c mice were randomized into 2 main groups, non-HLU and HLU groups. The mice from each group were further randomized into 4 subgroups receiving drinking water containing 0, 0.1, 1, and 10 g AHCC (provided by Amino Up Chemical Co, Ltd., Sapporo, Japan)/kg body weight/day. AHCC in powder form was weighed and dissolved in water at concentrations of 0, 0.4, 4 and 40 mg/ml for 0, 0.1, 1 and 10 g/kg body weight, respectively. These concentrations are based on the fact that mice usually drink 5 ml of water daily. All mice were fed the experimental drink and a commercial rodent diet ad libitum for the 7-days period of the experiment.
In the HLU group, the animals were subjected to HLU and housed in separate cages to simulate the isolation environment of microgravity [8, 16, 24]. Briefly, the tail of each mouse was cleaned with 70% isopropyl alcohol and air-dried. A piece of moleskin was cut to a size of 0.5 in x 2 in and used for tail suspension. The tip of a large paper clip was inserted through the moleskin strip. The moleskin was then wrapped around the tail in a coil-like manner at about one-third of the length from the base. To secure the moleskin in place, two small drops of nail glue adhesive were applied to both ends of the strip. The other end of the paper clip was unfolded and hooked onto a swivel-pulley system. The swivel pulley glides along a stainless steel rod that runs the length of the cage. The swivel provided the mouse with a full 360° range of movement. The angle of suspension was adjusted to about 20-25°, resulting in approximately 30% of weight resting on the forelimbs and allowing the mouse ample access to food and experimental drink. Mice were weighed before suspension and during experimental periods.
The PLN assay is based on the method described by Yamauchi et al. [8]. After the period of acclimatization, 10 BALB/c (H2d) and 10 C57BL/6 (H2b) mice were sacrificed by cervical dislocation. Spleens were removed aseptically and pooled in each group. To prepare single cell suspensions from the donor spleens (syngeneic BALB/c and allogeneic C57BL/6), the spleens were squeezed with two frost-sided glass slides in RPMI-1640 medium (Sigma, St. Louis, MO) containing 100 U/ml penicillin and 100 μg/ml streptomycin (Invitrogen Corporation, Grand Island, NY) and filtered through 50-mesh stainless steel screens. To remove red blood cells, the cells were treated with 0.83% ammonium chloride in 0.01 M Tris-HCl buffer (pH 7.5) (Sigma) at 37°C for 10 min. Then the cells were washed twice with RPMI-1640 medium and resuspended in RPMI-1640 medium. Cell concentration was adjusted to 2 x 108 cells/ml and then the cell suspension was irradiated (3000 R). The host BALB/c mice were injected into the hind footpad with 50 μl of 1 × 107 irradiated splenocytes from allogeneic C57BL/6 mice. The contralateral footpad was injected with 50 μl of 1 × 107 irradiated splenocytes from syngeneic BALB/c mice. After the inoculation, HLU treatment was started. On the seventh day of inoculation, the animals were sacrificed, and the PLNs were removed and weighed. The PLN immunoproliferative response to allogeneic challenge was calculated as the ratio of the allogeneic and syngeneic challenged PLN weights and reported as the stimulation index (SI).
After 7 days of AHCC feeding and HLU, the mice (different mice from PLN assay) were sacrificed. Their spleens were removed and weighed. Single-cell suspensions from spleens were prepared as described earlier. The splenocytes were resuspended in complete RPMI (RPMI-1640 medium supplemented with 10% heat-inactivated fetal bovine serum, 10 mM HEPES, 2 mM glutamine, 100 U/ml penicillin, and 100 μg/ml streptomycin (Invitrogen Corporation)). Cell concentration was adjusted to 2 x 106 cells/ml. Two × 105 splenocytes were stimulated with 2.5 μg/ml concanavalin A (Con A) (Biomeda Corp., Foster City, CA), in a 96-well flat-bottom plate in triplicate at 37°C in a humidified 5% CO2 incubator for 48 h. Cells cultured without mitogens served as unstimulated controls. For the last 16-18 h of culture, 1 μCi of [3H] thymidine deoxyribose (TdR) (GE Healthcare Bio-Sciences Corp., Piscataway, NJ) was added to the wells. Cultures were then harvested onto fiber-glass filters by using a PHD cell harvester (Cambridge Technology, Inc., Cambridge, MA). The filter disks were then transferred to 7 ml plastic disposable scintillation vials, and then 5 ml of liquid scintillation cocktail (MP Biomedicals, Solon, OH) was added. The amount of [3H ] TdR incorporated was measured in a Beckman LS-1800 liquid scintillation counter.
Splenocytes (1 × 106 cells/ml) in complete RPMI-1640 were cultured in a 24-well flat-bottom plate at 37°C in a humidified 5% CO2 incubator for 48 h with 5 μg/ml LPS. At the end of the incubation period, supernatants were collected and stored at -40°C until used. The cytokine production was quantified in duplicate with a Bio-Plex Suspension Array System (Bio-Rad Laboratories, Inc., Hercules, CA) according to the manufacturer's instructions.
Data were analyzed using two-way analysis of variance (ANOVA). Since there was no significant interaction between HLU and AHCC in all assays, one-way ANOVA was carried out within non- HLU and HLU, respectively followed by the scheffe post hoc test for multiple comparisons. Statistical differences between control and HLU groups were analyzed using the unpaired Student’s ttest. StatView software program (version 5.0; SAS Institute Inc., Cary, NC) was used. Data are expressed as means ± SEM. Differences were considered significant at p <0.05.
Results
AHCC supplementation increases body weight and spleen weight in the non- HLU group. However, we didn’t see the same effect in the HLU group with or without AHCC. Spleen weight, as an indicator of improved immune response, was higher with AHCC supplementation in both non-HLU and HLU groups (p < 0.05). After 7 days of HLU and AHCC supplementation, the increases in body weight and spleen weight were significantly smaller in mice in the HLU group than in mice in the non-HLU group (p< 0.001, p < 0.05, respectively) (Table I). AHCC dose of 1mg/kg seemed optimal in both on-HLU and HLU groups.
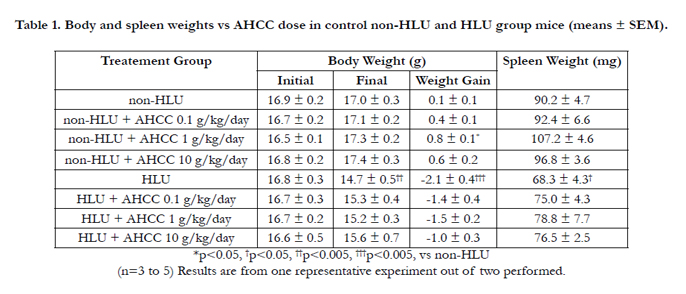

Table 1. Body and spleen weights vs AHCC dose in control non-HLU and HLU group mice (means ± SEM).
After 7 days of treatment, the PLN proliferative response to allogeneic challenge was examined to assess T cell function and immune response in vivo. The PLN proliferative response was significantly decreased in the HLU group compared to that in thenon-HLU group (p < 0.05). Supplementation with AHCC at doses of 1 and 10 g/kg/day significantly increased the PLN weight gain as immune response in the HLU group (p = 0.001 and p < 0.05, respectively) (Figure 1).

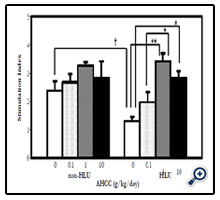
Figure 1. Stimulation Index for in vivo PLN proliferative response vs. AHCC dose in control and HLU mice (means ±SEM). *p <0.05, **p =0.001, †P <0.05 (n=3 to 5) Results are from one representative experiment out of two performed.
After 7 days of HLU and AHCC supplementation, we examined the in vitro splenocyte proliferative responses. T cell function was assessed by stimulation with mitogen Con A (concanavalin A) mitogen. There were no significant differences between the proliferative responses to Con A of splenocytes from mice in the non-HLU group and HLU group (Figure 2B). AHCC supplementation enhanced this response in a dose-dependent manner in the non-HLU group and was significant at 10 g/kg/day (p < 0.01). AHCC supplementation tended to increase this response in HLU group with 1g/kg/day dose showing higher response but shy of significance.
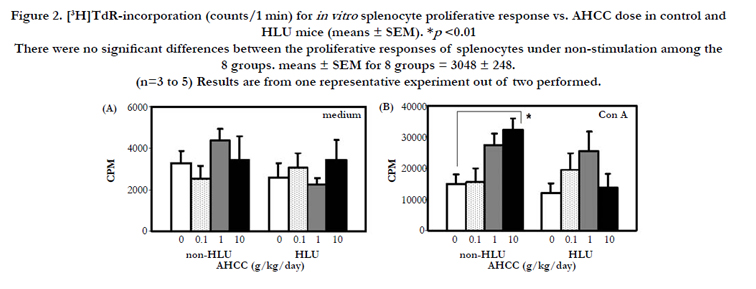
Figure 2. [3H]TdR-incorporation (counts/1 min) for in vitro splenocyte proliferative response vs. AHCC dose in control and HLU mice (means ± SEM). *p <0.01 There were no significant differences between the proliferative responses of splenocytes under non-stimulation among the 8 groups. means ± SEM for 8 groups = 3048 ± 248. (n=3 to 5) Results are from one representative experiment out of two performed.
We further examined in vitro cytokine and chemokine production from splenocytes. After stimulation of splenocytes with LPS (lipopolysaccharide), inflammatory cytokine and chemokine production was determined. The levels of IL-6, and IL-12 were significantly higher in the HLU group than in the non-HLU group(p <0.01 and p <0.05, respectively) (Figure 3C, D). The levels of IL-1α and IL-1β were higher in the HLU group than in the non- HLU group. Furthermore, the IL-1α level in mice in the AHCCfed HLU group was significantly higher than that in mice in the HLU group (p < 0.05) (Figure 3A, B). The IL-6 and IL-12 levels in the AHCC-fed HLU group tended to be higher than those in the HLU group.
Similar results were obtained for chemokine production. The levels of macrophage inflammatory protein (MIP)-1α, MIP-1α, granulocyte colony-stimulating factor (GCSF), and KC (murine homologue of IL-8) were significantly higher in the HLU group than in the non-HLU group (p < 0.001, p < 0.001, p < 0.005 and p < 0.01, respectively) (Figure 3E-H). The MIP-1α, MIP-1β, GCSF, and KC levels in mice in the AHCC-fed HLU group were higher than those in mice in the HLU group alone, but the differences were not significant.
Under normal physiological conditions with intact normal host immune response the inflammatory cytokine/chemokines should decrease. However, in HLU state physiological conditions, which exhibits similar physiological responses of space microgravity (which exhibits environmental effects) it is innate immune function is at risk so in this particular HLU analog presence of inflammatory mediators would be beneficial to protect subjects. Innate immune function comprises several mechanisms of humoral and phagocytic cell mediated responses and is first line of immune defense mechanism so in order to protect host.
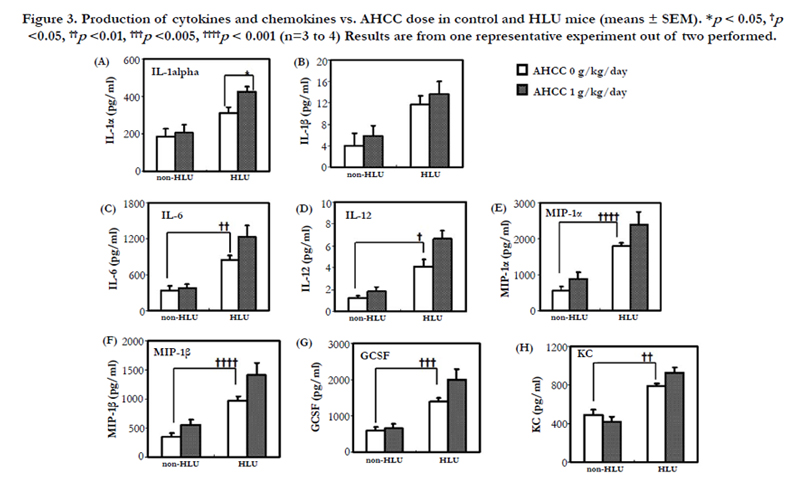
Figure 3. Production of cytokines and chemokines vs. AHCC dose in control and HLU mice (means ± SEM). *p < 0.05, †p<0.05, ††p <0.01, †††p <0.005, ††††p < 0.001 (n=3 to 4) Results are from one representative experiment out of two performed.
Discussion
In this study, we examined the effect of AHCC supplementation on microgravity-induced T cell dysfunction and found that AHCC supplementation has enhanced in vivo T cell function and tended to enhance in vitro T cell function. Our cytokine/chemokine study results showed that microgravity enhances inflammatory responses and AHCC tends to enhance those responses furthermore. There is definitely a need for countermeasures that will maintain normal immune system during spaceflight, especially when missions are prolonged. Since T cells play an important role in acquired immunity, a countermeasure for T cell dysfunction is imperative. A beneficial effect of AHCC on T cells has been reported [7, 21, 25]. We examined PLN response to allogeneic antigen as an in vivo immune response. PLN response is an in vivo cell-mediated immune response of hosts that involves all phases of immune response, e.g., antigen processing and presentation, followed by proliferative phase of immune response. PLN response was significantly decreased in mice in the HLU group compared to that in mice in the control group, and AHCC supplementation significantly reversed this response (Figure 1). These results suggest that AHCC reverses HLU-induced T cell dysfunction in PLNs. In vitro T cell function was assessed by stimulation with Con A (Figure 2B). There was no significant difference between proliferative responses to Con A in the control and HLU groups. Studies using HLU of rodents have shown that HLU produces organ and cell specific changes in the immune system rather than a generalized immunosuppression [6, 26]. Nash et al. reported that there were differences in lymphocyte response in HLU rats between peripheral blood lymphocytes (PBL), lymph nodes (LN) lymphocytes, and splenocytes. In HLU rats there was a significant decrease in proliferation in PBL, but only slight decreases in responses were observed in LN and splenocytes [26]. AHCC supplementation, as shown in Figure 1, enhanced PLN response in the HLU group. Response to Con A in the non-HLU group was slightly increased by AHCC administration but shy of significance. Our results suggest that efficacy of AHCC in T cell function in HLU model may differ among the tissues. In non-HLU, AHCC supplementation significantly enhanced splenocytes response to Con A (Figure 2). Aviles et al have shown that oral AHCC administration enhanced both Th1 and Th2 cytokine production from splenocytes in non- HLU and the production of Th1 cytokine was increased in HLU [7]. These finding suggests that AHCC does not completely reverse T cell function in HLU. Enhancement of T cell function is not for a specific T cell population but for overall T cells. Gao et al. reported that enhanced T cell function by AHCC was shown in CD4+ T cells, CD8+ T cells and γδT cells in the spleen and in draining lymph nodes in mice with tumors [21]. It has also been reported that oral AHCC supplementation suppressed thymic apoptosis induced by dexamethasone in rats [25]. Beneficial effects of AHCC for T cells may not be only in peripheral lymphoid tissue. The mechanism of augmentation of T cell function is unclear. However, it may not be a direct effect of AHCC on T cells. We cultured splenocytes from naive mice with AHCC supplementation in vitro and examined proliferation and Th1/Th2 cytokine production. Surprisingly, those responses were significantly decreased (unpublished observations). These results raise two possibilities. One is that metabolic compounds of AHCC may be related to increased immune function in vivo. The other is that AHCC may modulate some endocrine system related to T cell function. It has been reported that ferric nitrilotriacetate (FeNTA), which generates hydroxyl radicals, increased serum corticosterone level and that AHCC administration reduced a stress hormone modulating corticosterone level to the normal level [27]. We previously reported that serum level of corticosterone in HLU mice was significantly higher than that in non-HLU mice and that T cell response and proliferation in HLU mice was suppressed [8]. Therefore, elucidation of the effect of AHCC on the endocrine system will be important to understand the mechanism of T cell enhancement by AHCC in HLU mice.
On the other hand, innate immunity plays an important role at an earlier stage of infection. A substantial number of astronauts have alterations in their physiologic and immunohematologic response that would put them at risk for diseases during space flights [2]. Closed space and recycling of air and water may increase the risk of microbial load. Furthermore, humans usually carry opportunistic pathogens, latent bacteria and viruses, which may be reactivated and cause serious diseases under immune-compromised conditions, including spaceflight. Neutrophils-, macrophages-, or monocytes-mediated innate immunity is the first step to exclude pathogens. To assess overall functions of these types of cells, we measured the levels of LPS-stimulated cytokine and chemokine production from splenocytes. LPS-induced production of inflammatory cytokine (IL-1α, IL-1β, IL-6 and IL-12) and chemokine (MIP-1α, MIP-1β, GCSF and KC) was increased in mice in the HLU group compared to that in mice in the control group (Figure 3). In response to the proinflammatory responses in endotoxemia, host cells produce several anti-inflammatory cytokines, such as IL10 and IL-4, and inhibit the production of inflammatory cytokines (a). IL-10 is produced by Th2 cells, B cells and monocytes/ macrophages. The concomitant increase in IL-10 in HLU may be in response to increased proinflammatory cytokines (Figure 3). Augmented innate immune responses including inflammatory cytokine production, in an HLU group have been reported, and our finding is consistent with results of previous studies [24, 29, 30]. Increased innate immune responses have also been seen in actual spaceflights [31-33]. However, there have been studies showing decreased innate immune response caused by spaceflight [32, 33]. Therefore, the effects of HLU and spaceflight on innate immune response are not clear. It is likely that innate immune response was temporarily increased by HLU in our study. In our study, AHCC supplementation in HLU mice tended to further enhance the inflammatory cytokine and chemokine production (Figure 3). Aviles et al have shown that AHCC supplementation in HLU mice also increased IL-6 and TNF-α levels in peritoneal cells compared to the levels in HLU mice under the condition of stimulation with LPS and K. pneumoniae [7]. Aviles et al. also showed that AHCC supplementation before and during HLU prevented bacterial infection [10]. Those studies suggest that AHCC has an inflammatory effect. Our results and those of others suggest that enhancement of inflammatory cytokine and chemokine production by AHCC supplementation will be effective for preventing infection [7, 10]. To the best of our knowledge, this is the first study to show the effect of AHCC supplementation on inflammatory chemokine production.
The levels of IL-6, and IL-12 were significantly higher in the HLU group than in the non-HLU group (p < 0.01 and p < 0.05, respectively) (Figure 3C, D). The IL-6 and IL-12 levels in the AHCC-fed HLU group tended to be higher than those in the HLU group. Increased IL-12 (T cell stimulating factor) in the AHCC fed HLU mice indicates increased differentiation of naive T cells into Th1 cells. It also activates TNF-α and IFN-γ production by NK and T cells. Enhanced functional response can be demonstrated by IFN-γ production and killing of target cells [35]. Interleukin 6 (IL-6) is encoded by the IL-6 gene. IL-6 is secreted by T cells and macrophages to stimulate immune responses, e.g. during infection and after trauma, especially burns or other tissue damage leading to inflammation. IL-6 also plays a role in fighting infection, as IL-6 has been shown in mice to be required for resistance against bacterium Streptococcus pneumonia.
The levels of IL-1α and IL-1β were higher in the HLU group than in the non-HLU group. Furthermore, the IL-1α level in mice in the AHCC-fed HLU group was significantly higher than that in mice in the HLU group (p <0.05) (Figure 3A, B). These results show that at the end of 7 day HLU and 7 day HLU + AHCC, IL- 1α is significantly higher in the latter. IL-1α is a regulator of the immune response and can activate the TNF-α pathway. AHCC appears to enhance this response in HLU. Since IL-12 is also enhanced it appears that AHCC might act as an enhanced stimulator of the TNF-α pathway and increase adaptive immunity. This hypothesis needs to be elucidated in future experiments. The IL- 1β (inflammatory response mediator) levels should have been expected to lower in AHCC treated HLU mice since it is a COX 2 stimulator induces more inflammation (might not be suitable for long periods of time). However since these experiments were conducted with one week of HLU, perhaps the time window of cytokine synthesis is only adequate for observing an increased adaptive response. Also since this protein plays more of a role by induction of Cox-2 in the brain, the increase seen here may not be unusual.
The beneficial effect of AHCC supplementation found in this study is consistent and corroborative with results of previous studies using an HLU model [6, 10]. This is the first study to show that AHCC enhances T cell function in vivo in an HLU model. The mechanism of the immuno-enhancing effects of AHCC needs to be elucidated.
In summary, the results of this study suggest that AHCC supplementation enhances T cell immune function in microgravity and has a beneficial role in maintenance of the immune system.
Acknowledgments
This study was supported by a grant from Amino Up Chemical Company, Sapporo, Japan (ADK).
References
- Aponte VM, Finch DS, Klaus DM (2006) Considerations for non-invasive in-flight monitoring of astronaut immune status with potential use of MEMS and NEMS devices. Life Sci 79(14):1317-1333.
- Borchers AT, Keen CL, Gershwin ME (2002) Microgravity and immune responsiveness: Implications for space travel. Nutrition 18(10): 889-898.
- Sonnenfeld G (2005) The immune system in space, including Earth-based benefits of space-based research. Curr Pharm Biotechnol 6(4): 343-349.
- Morey-Holton ER, Globus RK (2002) Hindlimb unloading rodent model: technical aspects. J Appl Physiol 92(4):1367-1377.
- Chapes SK, Mastro AM, Sonnenfeld G, Berry WD (1993) Antiorthostatic suspension as a model for the effects of spaceflight on the immune system. J Leukoc Biol 54(3): 227-235.
- Armstrong JW, Nelson KA, Simske SJ, Luttges MW, Iandolo JJ, et al. (1993) Skeletal unloading causes organ-specific changes in immune cell responses. J Appl Physiol 75(6): 2734-2739.
- Aviles H, Belay T, Vance M, Sun B, Sonnenfeld G (2004) Active hexose correlated compound enhances the immune function of mice in the hindlimbunloading model of spaceflight conditions. J Appl Physiol 97(4):1437-1444.
- Yamauchi K, Hales NW, Robinson SM, Niehoff ML, Ramesh V, et al. (2002) Dietary nucleotides prevent decrease in cellular immunity in groundbased microgravity analog. J Appl Physiol 93(1): 161-166.
- Aviles H, Belay T, Fountain K, Vance M, Sonnenfeld G (2003) Increased susceptibility to Pseudomonas aeruginosa infection under hindlimb-unloading conditions. J Appl Physiol 95(1): 73-80.
- Aviles H, Belay T, Fountain K, Vance M, Sun B, Sonnenfeld G (2003) Active hexose correlated compound enhances resistance to Klebsiella pneumoniae infection in mice in the hindlimb-unloading model of spaceflight conditions. J Appl Physiol 95(2): 491-496.
- Belay T, Aviles H, Vance M, Fountain K, Sonnenfeld G (2002) Effects of the hindlimb-unloading model of spaceflight conditions on resistance of mice to infection with Klebsielia pneumoniae. J Allergy Clin Immunol 110(2):262-268.
- Lee EH, Ding W, Kulkarni AD, Granstein RD (2005) Tumor growth and immune function in mice during hind-limb unloading. Aviat Space Environ Med 76(6): 536-540.
- Taylor PW, Sommer AP (2005) Towards rational treatment of bacterial infections during extended space travel. Int J Antimicrob Agents 26(3): 183-187.
- Armstrong JW, Balch S, Chapes SK (1994) Interleukin-2 therapy reverses some immunosuppressive effects of skeletal unloading. J Appl Physiol 77(2):584-589.
- Chapes SK, Simske SJ, Forsman AD, Bateman TA, Zimmerman RJ (1999) Effects of space flight and IGF-1 on immune function. Adv Space Res 23(12): 1955-1964.
- Kulkarni AD, Yamauchi K, Hales NW, Ramesh V, Ramesh GT, et al. (2002) Nutrition beyond nutrition: plausibility of immunotrophic nutrition for space travel. Clin Nutr 21(3): 231-238.
- Kidd PM (2000) The use of mushroom glucans and proteoglycans in cancer treatment. Altern Med Rev 5(1): 4-27.
- Matsushita K, Kuramitsu Y, Ohiro Y, Obara M, Kobayashi M, et al. (1998) Combination therapy of active hexose correlated compound plus UFT significantly reduces the metastasis of rat mammary adenocarcinoma. Antican-cer Drugs 9(4): 343-350.
- Ritz BW, Nogusa S, Ackerman EA, Gardner EM (2006) Supplementation with active hexose correlated compound increases the innate immune response of young mice to primary influenza infection. J Nutr 136(11): 2868-2873.
- Aviles H, O'Donnell P, Sun B, Sonnenfeld G (2006) Active hexose correlated compound (AHCC) enhances resistance to infection in a mouse model of surgical wound infection. Surg Infect (Larchmt) 7(6): 527-535.
- Gao Y, Zhang D, Sun B, Fujii H, Kosuna K, et al. (2006) Active hexose correlated compound enhances tumor surveillance through regulating both innate and adaptive immune responses. Cancer Immunol Immunother 55(10): 1258-1266.
- Daddaoua A, Martinez-Plata E, Lopez-Posadas R, Vieites JM, Gonzalez M, et al. (2007) Active hexose correlated compound acts as a prebiotic and is anti inflammatory in rats with hapten-induced colitis. J Nutr 137(5): 1222-1228.
- Cowawintaweewat S, Manoromana S, Sriplung H, Khuhaprema T, Tongtawe P, et al. (2006) Prognostic improvement of patients with advanced liver cancer after active hexose correlated compound (AHCC) treatment. Asian Pac J Allergy Immunol 24(1): 33-45.
- Felix K, Wise K, Manna S, Yamauchi K, Wilson BL, et al. (2004) Altered cytokine expression in tissues of mice subjected to simulated microgravity. Mol Cell Biochem 266(1-2): 79-85.
- Burikhanov RB, Wakame K, Igarashi Y, Wang S, Matsuzaki S (2000) Suppressive effect of active hexose correlated compound (AHCC) on thymic apoptosis induced by dexamethasone in the rat. Endocr Regul 34(4): 181-188.
- Nash PV, Bour BA, Mastro AM (1991) Effect of hindlimb suspension simulation of microgravity on in vitro immunological responses. Exp Cell Res195(2): 353-360.
- Ye SF, Wakame K, Ichimura K, Matsuzaki S (2004) Amelioration by active hexose correlated compound of endocrine disturbances induced by oxidative stress in the rat. Endocr Regul 38(1): 7-13.
- Calcagni E, Elenkov I (2006) Stress system activity, innate and T helper cytokines, and susceptibility to immune-related diseases. Ann N Y Acad Sci 1069: 62-76.
- Miller ES, Bates RA, Koebel DA, Sonnenfeld G (1999) Antiorthostatic suspension stimulates profiles of macrophage activation in mice. Neuroimmunomodulation6(3): 160-167.
- Wen XL, Yang GH, Wang T, Hu P (2001) Effects of simulated weightlessness on T cell subpopulations and activity of IL-2 and IL-6 in mice. Space Med Med Eng (Beijing) 14(1): 60-62.
- Chapes SK, Morrison DR, Guikema JA, Lewis ML, Spooner BS (1994) Production and action of cytokines in space. Adv Space Res 14(8): 5-9.
- Kaur I, Simons ER, Castro VA, Mark Ott C, Pierson DL (2004) Changes in neutrophil functions in astronauts. Brain Behav Immun 18(5): 443-450.
- Stowe RP, Sams CF, Mehta SK, Kaur I, Jones ML, et al. (1999) Leukocyte subsets and neutrophil function after short-term spaceflight. J Leukoc Biol.; 65(2): 179-186.
- Steffen JM, Musacchia XJ (1986) Thymic involution in the suspended rat: adrenal hypertrophy and glucocorticoid receptor content. Aviat Space Environ Med 57(2): 162-167.
- Hsieh CS, Macatonia SE, Tripp CS, Wolf SF, O'Garra A, et al. (1993) Development of TH1 CD4+ T cells through IL-12 produced by Listeriainduced macrophages. Science 260(5107): 547-549.
- van der Poll T, Keogh CV, Guirao X, Buurman WA, Kopf M, et al. (1997) Interleukin-6 gene-deficient mice show impaired defense against pneumococcal pneumonia. J Infect Dis 176(2): 439-444.
- Bankers-Fulbright JL, Kalli KR, McKean DJ (1996) Interleukin-1 signal transduction. Life Sci 59(2): 61-83.
- Dinarello CA (1997) Induction of interleukin-1 and interleukin-1 receptor antagonist. Semin Oncol 24(3 Suppl 9): S9-81-S9-93.
- Kim SH, Mok JW, Kim HS, Joo CK (2008) Association of -31T>C and -511 C>T polymorphisms in the interleukin 1 beta (IL1B) promoter in Korean keratoconus patients. Mol Vis 14: 2109-2116.





