An Investigation of the Visual Function of Dyslexic Children
Ostadimoghaddam H1,2, Sobhani Rad D3, Esmaeili A4, Salehinejad Z4*, Ehsaei A1,2, Heravian S J1,2, Yekta AA1,2
1 Refractive Errors Research Center, School of Paramedical Sciences, Mashhad University of Medical Sciences, Mashhad, Iran.
2 Department of Optometry, School of Paramedical Sciences, Mashhad University of Medical Sciences, Mashhad, Iran.
3 Department of Speech Therapy, School of Rehabilitation, Mashhad University of Medical Sciences, Mashhad, Iran.
4 Department of Ophthalmology, Razavi Hospital, Mashhad, Iran.
5 Student Research Committee, Mashhad University of Medical Sciences,Mashhad, Iran.
*Corresponding Author
Zeinab Salehinejad MSc.,
Student Research Committee,
Department of Optometry,
Mashhad University of Medical Sciences,
Mashhad, Iran.
Tel: 00989153089552
E-mail: zeinabsalehi@rocketmail.com
Article Type: Research Article
Received: June 01, 2015; Accepted: June 09, 2015; Published: June 10, 2015
Citation: Salehinejad Z et al., (2015) An Investigation of the Visual Function of Dyslexic Children. Int J Ophthalmol Eye Res 03(5), 110-113. doi: dx.doi.org/10.19070/2332-290X-1500022
Copyright: Zeinab S© 2015. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
Background: Dyslexia is the most common learning disability, accounts for 5 to 17% of the school-aged children. Dyslexia is defined as an impairment in which the child faces difficulties learning, reading and writing relative to IQ.
Objective: The aim of this study was to investigate the central and peripheral visual function in a group of dyslexic children who write from right to left direction.
Materials/Patients and Methods: Twenty dyslexic eyes and 20 normal eyes were examined in this study. The visual examinations consisted of measuring distant and near visual acuity, refraction and visual field evaluation using an Octopus 101 automated perimeter. Foveal sensitivity was also measured with static perimetry, and then kinetic perimetry in two target brightness levels: 15 and 20 dB and data were analyzed by SPSS 17 statistical software.
Results: Dyslexic and normal children of the study population aged from 7 to 10 years with mean age of 8.5. Mean visible surface area at brightness levels of 15 and 20 dB was significantly different between two groups (p<0.001, p=0.001). In addition, inferior hemifield had the most noticeable differences between groups compared to other hemifields. No significant difference was noted between two groups for other examinations such as visual acuity at distance and near, refractive errors and static foveal sensitivity.
Conclusions: Visible surface area was defected in dyslexic children; however, the foveal function was similar to normal individuals. These results support the hypothesis of an impairment of the magnocellular function in dyslexia, whereas the parvocellular pathway is intact.
2.Background
3.Objective
4.Materials/Patients and Methods
5.Results
6.Discussion
7.Acknowledgements
8.References
Keywords
Developmental Dyslexia; Kinetic Perimetry; Foveal Sensitivity; Parvocellular Pathway; Magnocellular pathway.
Background
Dyslexia is the most common learning disability, accounts for 5 to 17% of the school-aged children [1, 2]. Dyslexia is defined as an impairment in which the child faces difficulties learning, reading and writing relative to IQ [3]. This developmental disorder affects lots of acquired skills in addition to reading, writing and spelling, such as skills automatisation, balance, phonological awareness, sequential tasks, handwriting and articulation. It has been proved that all these deficits are caused by a disorder in cerebellum [4].
It has been suggested that deficit in sequencing and visual information processing are caused by a disorder in magnocellular pathway [3]. In addition, cerebellum is the head ganglion of magnocellular system [5]. Therefore, it may be hypothesised that all the observed difficulties in dyslexics are caused by the congenital disorder in cerebellum. Majority of previous investigations have focused on developmental disorders including dyslexia. Since dyslexia is usually detected when reading started in school-aged children, and considering visual system role in reading, investigators have always interested in finding the relationship between the dyslexia and vision. Different aspects of the visual function have been extensively investigated in dyslexia such as routine visual acuity tests [6-8] and more complex tests like visual evoked potential and electroretinogram [9, 10]. However, controversies exist regarding the outcomes of the above-mentioned studies.
Objective
Considering the most reliable theory of the etiology of dyslexia, impairment in the visual magnocellular system causes reading problems [11-13]. Visual magnocellular system is activated with peripheral stimulus, low-spatial frequency, fast movement or changes in speed [14]. Contrast sensitivity [15], visual evoked potential [9,10] random dots kinetogram and flicker sensitivity tests [15] have been used extensively to evaluate visual magnocelluar system, however, results of the majority of these investigations are not well-matched with this theory [16-18] Kinetic perimetry is another test which evaluates the magnocelluar system with moving target. This test has not been fully evaluated in dyslexia and the results of previous reported studies are not similar [19-21].
Furthermore, reading habit (from left to right or from right to left) is an important factor in reading skills. However, the number of studies which have been performed in countries, in which reading habits are from right to left (e.g. Iran), is very limited. [22] The present study has evaluated the ability of detection acuity with kinetic perimetry in dyslexic school-aged children and compared the results with normal matched control group. Static perimetry and other routine visual examinations such as visual acuity and refraction have been also performed.
Materials/Patients and Methods
Ten dyslexic school-aged children and 10 normal age, gendermatched children (total of 40 eyes) were recruited for this casecontrol study. Informed consent was obtained from each participant (or the patient’s parent or legal guardian), after the nature of the experimental procedures had been explained. The research followed the tenets of the Declaration of Helsinki and was approved by Mashhad University of Medical Sciences Research Ethics Committee.
In this study, evaluation was performed by speech therapist and based on the available history and records to detect children with dyslexia. Participants with other physical problems (e.g. hearing difficulty) were excluded from the study population. In addition, children's IQ was tested with Wechsler test and those with any problems were excluded from the study population. Afterwards, visual examinations were performed in an eye clinic. Visual acuity was measured with Snellen chart using appropriate destination and room lighting condition.
In addition, vision was evaluated without correction and with habitual correction. Near visual acuity was also tested with Snellen near chart. To ensure that the subjects met the inclusion criteria, their sphero-cylindrical refractive errors were measured by subjective refraction collected by a qualified optometrist. As the first step for each participant and to determine the refraction, a routine non-cycloplegic subjective refraction was performed. All refractions were performed by the same examiner. The traditional endpoint of maximum plus/minimum minus to achieve the optimum visual acuity was adopted. To determine astigmatism, a crossed cylinder was used to locate the axis and power. All cases had visual acuity of 6/6 or better. In addition, fundoscopy was performed with direct ophthalmoscope and children with any posterior segment disorder were excluded.
Octopus perimeter 101 (Octopus 101, Haag-Streit Inc., Koeniz, Switzerland) was used for assessing static and kinetic visual field. Initially, static preimetry for central two degrees of the macula was performed. Foveal threshold contrast was obtained based on the presentation of 20 small bright light points (with 4 mm diameter). Then, kinetic perimetry, with a bright light moving target (1mm diameter and 4 degree per second speed), was performed in different axis. Measured locations included 0, 45, 90, 135, 180, 225, 270 and 335 degree eccentricities. The subjects were asked to look at the central fixing point, hold a button in his hands and press it as soon as he/she sees the moving stimulus in peripheral visual field. These pointes were recorded automatically relative to the center. Target was presented in two different brightness 15 and 20 db. Visual field experiment was performed for the right eye and the left eye, respectively. Isopters were drawn and total area for each eye in each brightness, and area for each hemifields (right, left, up and down) were recorded. These areas are presented in kinetic perimetry with degree2 and then it is transformed according to the following formula to ease comparison:
R is the perimeter hemisphere diameter (in centimeter), and deg2 is calculated area based on the degree power which is presented by the system after drawing isopters. Statistical analysis was performed using SPSS version 17 (SPSS Inc., Chicago, Illinois, USA). Kolmogorov-Smirnov test was used to ascertain the normality of the data and in the case of normal distribution; t test was used to compare the results between groups and genders.
Results
20 children aged from 7 to 10 years with mean age of 8.5 participated in the current study. Our results showed that the mean spherical equivalent (MSE) were not significantly different between two groups (Table 1).
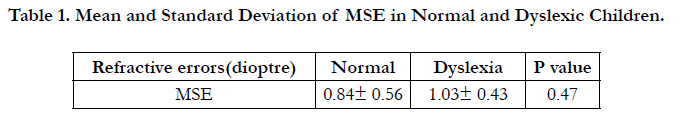

Table 1. Mean and Standard Deviation of MSE in Normal and Dyslexic Children.
The average of foveal sensitivity was not also significantly different between two groups (Table 2).

Table 2. Mean and Standard Deviation of Foveal Sensitivity in Normal and Dyslexic Children.
However, kinetic visual field results showed that the mean of visible surface area in two brightness levels was significantly different between normal and dyslexic children and the smallest p-value belongs to the brightness of 15 db (Table 3).
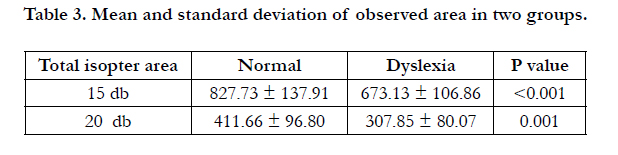
Table 3. Mean and standard deviation of observed area in two groups.
Visible surface area of each hemifield isopter was also compared between groups and in all cases the difference was statistically significant. The smallest p-value belongs to the inferior hemifield (Table 4).
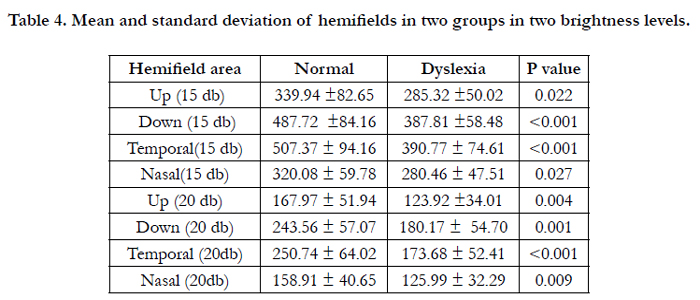
Table 4. Mean and standard deviation of hemifields in two groups in two brightness levels.
Discussion
The present study investigated the visual function between schoolaged dyslexic and normal children. Based on the present results, the average of refractive errors were not significantly different between dyslexic and normal groups which is similar to the results of Latvala and Dusek [7, 8]. The mean of the foveal sensitivity was not significantly different between two groups, similar to the previous reported findings. Stevens and Neville showed that parvocellular system function in dyslexic individuals is not different from normal population. These results confirm the magnocellular system defect theories [19]. Grosser and Spafford found significant differences in static perimetry test results between dyslexic group and a group with high reading ability. Disparity between our findings and their results could be due to including individuals with high reading ability in their control group [21].
Kinetic visual field results in the present study showed that the total area in both measured brightness levels and for both eyes were significantly different between dyslexic and normal children; Stevens found similar results [19]. The inferior hemifields showed the most obvious differences between two groups which is complied with the magnocellular defect theory.
Based on other investigations inferior part of the visual field is mostly presented by the magnocellular pathway. This may explain the most significant differences between two groups in inferior hemifields and is similar to previous studies [19, 23].
Acknowledgements
We thank the research vice chancellor of Mashhad University of Medical Sciences for supporting this study (grant number: 89311). The results described in this paper were part of a thesis for a Master degree in Optometry and Vision Science.
References
- Shaywitz S (1998) The New England journal of medicine. Dyslexia 338(5): 307-312.
- Sun Z, Zou L, Zhang J, Mo S, Shao S, Zhong R, et al. (2013) Prevalence and associated risk factors of dyslexic children in a middle-sized city of China: a cross-sectional study. PloS one.8(2): e56688.
- Demb JB, Boynton GM, Best M, Heeger DJ (1998) Psychophysical evidence for a magnocellular pathway deficit in dyslexia. Vision research. 38(11): 1555-1559.
- Evans BJ, Drasdo N (1990) Review of ophthalmic factors in dyslexia. Ophthalmic Physiol Opt. 10(2): 123-132.
- Stein J (2001) The magnocellular theory of developmental dyslexia. Dyslexia. 7(1): 12-36.
- Evans BJ, Drasdo N, Richards IL (1994) An investigation of some sensory and refractive visual factors in dyslexia. Vision research. 34(14): 1913-26.
- Latvala ML, Korhonen TT, Penttinen M, Laippala P (1994) Ophthalmic findings in dyslexic schoolchildren. The British journal of ophthalmology. 78(5): 339-343.
- Dusek W, Pierscionek BK, McClelland JF (2010) A survey of visual function in an Austrian population of school-age children with reading and writing difficulties. BMC ophthalmology.10-16.
- Livingstone MS, Rosen GD, Drislane FW, Galaburda AM (1991) Physiological and anatomical evidence for a magnocellular defect in developmental dyslexia. Proceedings of the National Academy of Sciences of the United States of America. 88(18): 7943-7.
- Romani A, Conte S, Callieco R, Bergamaschi R, Versino M, Lanzi G, et al. (2001) Visual evoked potential abnormalities in dyslexic children. Functional neurology. 16(3): 219-29.
- Talcott JB, Hansen PC, Assoku EL, Stein JF (2000) Visual motion sensitivity in dyslexia: evidence for temporal and energy integration deficits. Neuropsychologia. 38(7): 935-43.
- Galaburda AM, Menard MT, Rosen GD (1994) Evidence for aberrant auditory anatomy in developmental dyslexia. Proceedings of the National Academy of Sciences of the United States of America. 91(17): 8010-8013.
- Borsting E, Ridder WH 3rd, Dudeck K, Kelley C, Matsui L, Motoyama J (1996) The presence of a magnocellular defect depends on the type of dyslexia. Vision research. 36(7): 1047-53.
- Zeki S, Watson JD, Lueck CJ, Friston KJ, Kennard C, Frackowiak RS (1991) A direct demonstration of functional specialization in human visual cortex. The Journal of neuroscience : The official Journal of the Society for Neuroscience. 11(3): 641-649.
- Martin F, Lovegrove W (1987) Flicker contrast sensitivity in normal and specifically disabled readers. Perception. 16(2): 215-21.
- Graves RE, Frerichs RJ, Cook JA (1999 ) Visual localization in dyslexia. Neuropsychology. 13(4): 575-81.
- Skottun BC (2000) The magnocellular deficit theory of dyslexia: The evidence from contrast sensitivity. Vision research. 40(1): 111-27.
- Williams MJ, Stuart GW, Castles A, McAnally KI (2003) Contrast sensitivity in subgroups of developmental dyslexia. Vision research. 43(4): 467-77.
- Stevens C, Neville H (2006 ) Neuroplasticity as a double-edged sword: deaf enhancements and dyslexic deficits in motion processing. Journal of cognitive neuroscience. 18(5): 701-14.
- Grosser GS, Spafford CS (1989) Perceptual evidence for an anomalous distribution of rods and cones in the retinas of dyslexics: a new hypothesis. Perceptual and motor skills. 68(3 Pt 1): 683-98.
- Spafford C, Grosser GS (1991) Retinal differences in light sensitivity between dyslexic and proficient reading children: new prospects for optometric input in diagnosing dyslexia. Journal of the American Optometric Association. 62(8): 610-5.
- Farrag AF, Khedr EM, Abel-Naser W (2002) Impaired parvocellular pathway in dyslexic children. European journal of neurology : the official journal of the European Federation of Neurological Societies. 9(4): 359-63.
- Previc FH (1990) Functional specialization in the lower and upper visual fields in humans: Its ecological origins and neurophysiological implications. Behavioral and Brain Sciences. 13(03): 519-42.





