Double Stranded RNA Simultaneously Targeting Four White Spot Syndrome Virus (WSSV) genes Provides Protection against WSSV in Litopenaeus Vannamei
Anil Kumar1,2*, Laramore S3, Alexander P4, F. C. Thomas Allnutt5, Sayre RT1,2
1 Donald Danforth Plant Science Center 975 N. Warson Rd, St. Louis, MO 63132, USA.
2 New Mexico Consortium, Los Alamos National Laboratory, 4200 W Jemez Rd., Los Alamos, NM 87544, USA.
3 Harbor Branch Oceanographic Institute at Florida Atlantic University, 5600 US 1 North, Fort Pierce, FL, 34946, USA.
4 Logos Energy, 51 Alpha Park, Highland Heights, Ohio 44143, USA.
5 Phycal Inc., 51 Alpha Park, Highland Heights, OH 44143, USA.
*Corresponding Author
Anil Kumar,
Donald Danforth Plant Science Center 975 N. Warson Rd,
St. Louis, MO 63132, USA.
E-mail: anil.kumar@gmail.com
Received: March 03, 2015; Accepted: April 08, 2015; Published: April 13, 2015
Citation: Anil Kumar, et al., (2015) Double Stranded RNA Simultaneously Targeting four White Spot Syndrome Virus (WSSV) genes provides protection against WSSV in Litopenaeus Vannamei. Int J Marine Sci Ocean Technol. 2(2), 5-10.
Copyright: Anil Kumar© 2015. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
White Spot Syndrome Virus (WSSV) is amajor economic threat to global shrimp aquaculture for which there is no available therapeutic. RNA-interference (RNAi)-based strategies could be a promising approach for limiting or curing WSSV epizootics. The present study explored the duration of effective protection provided to shrimp following treatment with long dsRNA molecules targeting WSSV genes. Injection of a mixture of in vitro transcribed dsRNA targeting four WSSV genes (viral capsid protein 28 (vp28), viral capsid protein 19 (vp19), ribonucleotide reductase2 (rr2) and thymidine kinasethymidylate kinase (tk-tmk) provided protection up to 95% for treated shrimp for up to 28 days post-injection. Periodic injection of 2.0 μg of the dsRNA mixture was as effective as injection of a single 15 μg dose in protecting shrimp against WSSV. These data have significant practical implications by providing further knowledge for development of RNAi-based therapeutics against WSSV.
2.Introduction
3.Materials and Methods
3.1. dsRNA Synthesis
3.2. Shrimp Injection Trial
3.3. Polymerase Chain Reaction (PCR) Analysis
4.Results
4.1. Injection of dsRNA targeting four WSSV genes provided effective protection against WSSV up to 28 days post injection
4.2. WSSV was not detected in shrimp injected with dsRNA targeting WSSV genes
5.Discussion
6.Conclusion
7.References
Keywords
WSSV; RNAi; Shrimp; Aquaculture.
Introduction
Shrimp aquaculture suffers significant annual losses on a global basis due to viral disease outbreaks. White Spot Disease (WSD), caused by the White Spot Syndrome Virus (WSSV), has been the most problematic viral pathogen affecting global shrimp farming since its emergence in 1992. WSSV is highly virulent and may result in 80-100% mortality in ponds within 7-10 days of infection [13]. The cumulative economic losses due to WSSV epidemics to the shrimp aquaculture industry stand at over 8 billion dollars since its first appearance in 1992 [5].
WSSV is one of the most widespread viruses in the shrimp industry, occurring in virtually all shrimp farming countries. Over 100 crustacean species have been found to be susceptible to WSSV, including shrimp, crayfish, crabs, and lobsters, these hosts can also act as carriers of the virus [1] The two most widely cultivated species of shrimp, Litopenaeus vannamei (Pacific white shrimp) and Penaeus Monodon (tiger shrimp) are highly susceptible to WSSV infection. WSSV is a very large bacilliform, non-occluded, enveloped virus with a flagella-like appendage at one end of the viral particle [3, 24, 27, 6]. The WSSV genome is one of the largest viral genomes sequenced, consisting of a circular, double-stranded DNA molecule ranging from 290 to 315 kb, depending upon the isolate [23, 31]. The genome is predicted to contain about 181 protein encoding open reading frames [23, 31, 6].
Various strategies have been tried to control WSSV infection in shrimp, including use of inactivated virus and recombinant protein- based subunit vaccines [18, 26, 11]. To date, no validated vaccine or therapeutic agent against WSSV is available in the market. RNA interference (RNAi)-based strategies appear tobe promising for the control of WSSV and other viral diseases of shrimp [20]. RNAi refers to a cellular mechanism that is initiated via double stranded RNA (dsRNA) resulting in inhibition of gene expression through chromatin modification, translational repression or sequence-specific degradation of target RNA [7, 16]. The RNAi machinery appears to be conserved among most eukaryotes, including invertebrates, and involves three steps: (i) processing of long dsRNA into 21 to 23 nucleotide fragments called small interfering RNAs (siRNAs) by an RNase III family enzyme called Dicer (Hammond et al. 2000), (ii) incorporation of siRNAs into the RNA-induced silencing complex (RISC), and (iii) siRNA-mediated guiding and cleavage of target messenger RNA (mRNA) by the RISC resulting in gene silencing [8, 9, 15]. Novel biotechnological applications of RNAi such as animal gene therapy [4, 22], control of mosquitoes [12] andcontrol of insect-pests of important agricultural crops [10, 30, 2] are being explored. The potential of RNAi for controlling the replication and spread of WSSV has also been explored. Table 1 provides a summary of studies conducted on evaluating the effectiveness of dsRNA treatment on protecting shrimp against WSSV infection. The first report demonstrating that exposure of marine shrimp to dsRNA induces innate antiviral immunity in a sequence independent manner was published in 2004 [19]. Since then, several studies demonstrated that injection of dsRNA targeting ribonucleotide reductase small subunit (rr2), DNA polymerase (dnapol), viral protein 28 (vp28) and viral protein 19 (vp19) provides significant and potent antiviral immunity against WSSV compared to injection of non-specific dsRNA [20, 17]. Oral delivery of bacterially-expressed vp28 dsRNA has also been reported to provide some protection [21]. While, most of these studies involved challenge of treated shrimp with the virus within 72 h post-injection of RNAi elements [20, 25, 29], only one study tried to evaluate the effectiveness of RNAi treatment in protecting shrimp against WSSV 10 days and 20 days post-treatment [17]. For anRNAi-based treatment to be commercially viable, it should be able to provide protection against WSSV for at least 7 to 10 days post-RNAi treatment. Otherwise, the cost of delivering RNAi elements, either through feed or injection, would make it impossible for shrimp farmers to adopt this technology. The present study developed novel RNAi elements targeting WSSV, tested their abilityto control WSSV by injection, and demonstrated the efficacy of the RNAi-elements to provide effective protection for 28 days post-delivery of RNAi elements to Litopenaeus vannamei against WSSV.
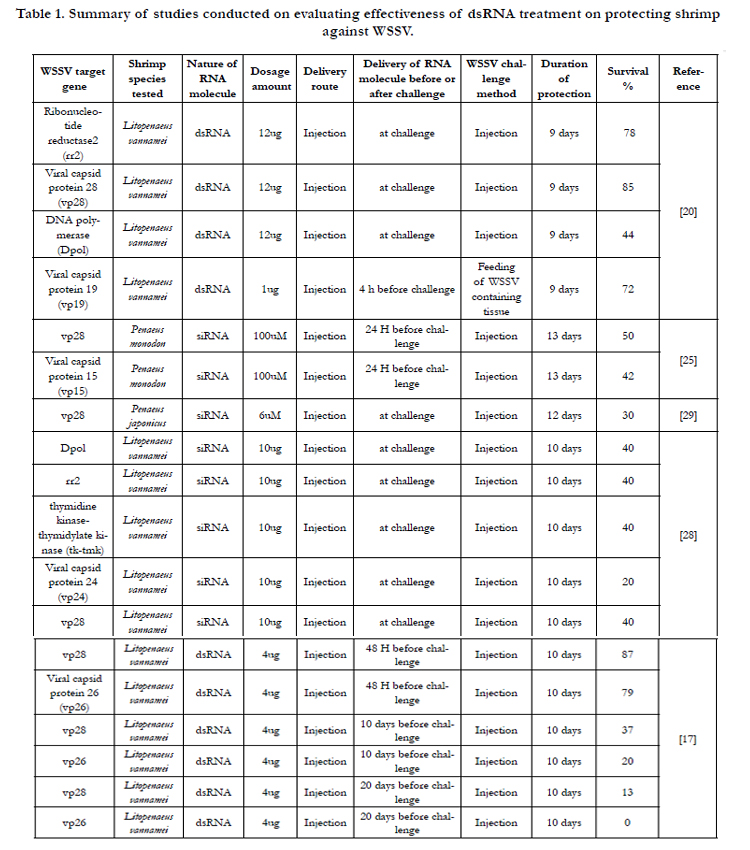

Table 1. Summary of studies conducted on evaluating effectiveness of dsRNA treatment on protecting shrimp against WSSV.
Materials and Methods
Two DNA fragments, one consisting of 273 bp of vp28 (GenBank Accession No. HM484390.1) and 266 bp of vp19 (GenBank Accession No. AY316119.1) and another consisting of b 251 bp of the rr2 (GenBank Accession No. AF144620.1) and 260 bp of the tk-tmk (GenBank Accession No. FJ756475.1) genes of WSSV were created by DNA synthesis (Epoch Life Science, Texas, USA). PCR reactions were set with primers T7- vp28 (5’-ATTAATACGA CTCACTATAG GGAGACACAG GCAATATCGA GACAAACATG GATGAAAAC-3’) and T7- vp19 (5’-GATAATACGA CTCACTATAG GGAGATTATA TTTGTCCTCA TCATCGTTAT CGTTGGC -3’) to add the T7 promoter to both ends of the vp28-vp19 fusion. The PCR product from the above reaction was used as a template to synthesize vp28-vp19 dsRNA using MEGA script RNAi kit (Life Technologies, Carlsbad, California, USA) following the manufacturer’s protocol. Similarly, PCR reactions were setwith the primers T7-rr2 (5’-ATTAATACGACTCACTATAGGGAGAGAGTGGGCTATTAATTGGATGCAAAG-3’) and T7-tk-tmk (5’-GATAATACGACTCACTATAGGGAGATTAGATGCTAGCTTAACAATGGAATTTGTATAAG-3’) to add the T7 promoter to both ends of rr2-tk-tmk fusion. The PCR product was then used as the template to synthesize rr2-tk-tmk dsRNA using the-MEGA script RNAi kit (Life Technologies, Carlsbad, CA) following the manufacturer’s protocol. Quality and concentration of vp28-vp19 and rr2-tk-tmk dsRNAs was measured using a Nanodrop® spectrophotometer (Thermo Fisher Scientific Inc., Waltham, Maryland, USA). A small aliquot of dsRNA from each of the two reactions was run on a 1% agarose gel to check the quality and size of dsRNA molecules.
Equal amounts of vp28-vp19 and rr2-tk-tmk dsRNAs were pooled to create a master mix dsRNA for injection. Juvenile SPF (specific pathogen free) Litopenaeus vannamei (these are at approximately post-larval stage 45 (~PL45) and at 0.5-1.0 g initial weight) obtained from Shrimp Improvement Systems (Islamorada, FL) were used for the study. Twelve shrimp were placed in each of the 18 x 38 L tanks, which were each provided with filtration, containing UV treated saltwater (32 ppt) and maintained at 25°C. Shrimp were acclimated to tank conditions for three days prior to the start of the experiment. Water quality parameters (temperature, salinity, dissolved oxygen, and pH) were recorded daily and ammonia and nitrite levels tested weekly. Water changes of 25% were conducted on every other day. Dissolved oxygen, salinity, pH, ammonia and nitrite levels remained in the normal range during the entire duration of the study and no differences were seen with regard to tank position in the test system. Shrimp were assigned to one of six groups: single or multiple dose treatments challenged at 5 or 14 days, positive or negative control (see Table 2) and were administered dsRNA (one 15 μg or three 2 μg doses in 20 μL of saline) or 20 μL saline (controls) via intramuscular injection, three replicates per treatment group. All treatment groups and the positive control group were WSSV challenged per os with minced tissue from freshly dead, WSSV-infected shrimp at either 5 or 14 days following treatment with dsRNA. The negative control group was not challenged. Mortality was followed and recorded for 14 days post-challenge.

Table2. Description of dsRNA injection treatments tested in this study.
Dead (frozen, collected) and surviving shrimp were analyzed for the presence of WSSV following experiment termination.Total shrimp genomic DNA was purified from 25-50 mg tissue samples using the Wizard Genomic DNA Purification Kit (Promega Corp., Madison, WI) according to the manufacturer’s instructions. Presence of WSSV in tissue samples was confirmed by using the primer pair 146F/R [14] using pooled DNA extracted from samples consisting of no more than 5 individuals per tank. Purified DNA was amplified in a 50 μL sample volume containing Choice-Taq (1 μL), a 10x buffer (5.0 μL), 25 mM dNTP’s (1μL), 1 μL of the appropriate forward and reverse primer (20 μM), 1 μL of template DNA and 40.0μL of sterile nuclease free water in a BioRad “iCycler” thermocycler. PCR conditions were as follows: 95°C for 30 s, followed by 35 cycles of 94°C for 30 s, an annealing temperature of 60°C for 30 s and an extension temperature of 72°C for 1 min, followed by a final extension of 72°C for 5 min.
Four WSSV genes, vp28, vp19, rr2 and tk-tmk were targeted for control of the WSSV in shrimp. The dsRNA targeting these four genes was synthesized by in vitro transcription. Four treatments were used to test the efficacy of RNAi in limiting WSSV in shrimp. The treatments listed in Table 2 were designed to assess the dosage required to provide effective protection and to evaluatethe duration of effective protection provided by the RNAi elements against WSSV.
Treatments T1 and T2, 15 μg single dose and three 2 μg doses challenged after 5 d, respectively, were both effective in providing protection to treated shrimp. While 80% of the shrimp survived in T1, the survival in T2 was 100% (Figure 1A). With the negative control (untreated shrimp not challenged with WSSV) 94% survival was observed. With the positive control (untreated shrimp challenged with WSSV) 42% mortality was observed two days post-challenge, which increased to 80% on the third day and reached 100% mortality on the fifth day (Figure 1A).
Treatments T3 and T4, 15 μg single dose and three 2 μg doses challenged after 14 d, respectively, also provided effective protection to treated shrimp. Survival percentage of 94% was observed with both treatments (Figure 1B). The results of T3 and T4 are significant as they indicated that RNAi elements can survive and remain active in shrimp for 14 or more days and multiple small dosages of dsRNA (T4) were as effective as a single large dosage of dsRNA (T3) in protecting shrimp for 28 days. Figure 1C summarizes the results of all the four treatments tested.
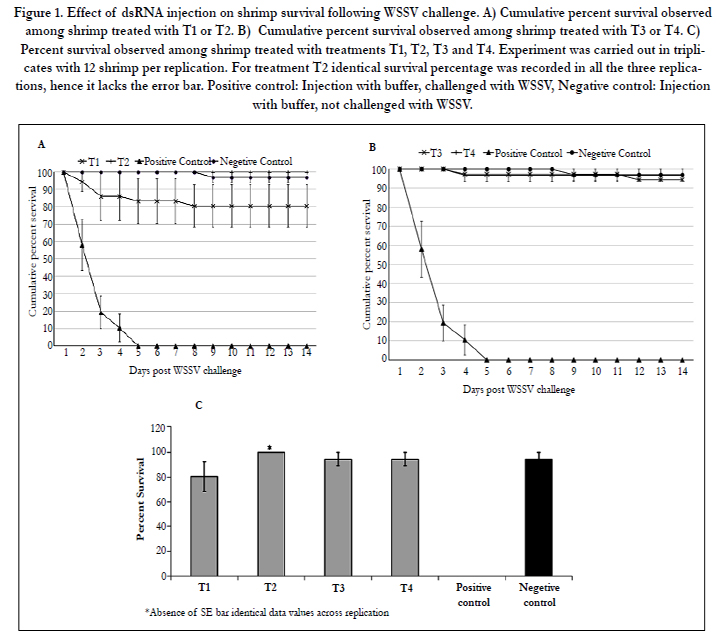
Figure 1. Effect of dsRNA injection on shrimp survival following WSSV challenge. A) Cumulative percent survival observed among shrimp treated with T1 or T2. B) Cumulative percent survival observed among shrimp treated with T3 or T4. C) Percent survival observed among shrimp treated with treatments T1, T2, T3 and T4. Experiment was carried out in triplicates with 12 shrimp per replication. For treatment T2 identical survival percentage was recorded in all the three replications, hence it lacks the error bar. Positive control: Injection with buffer, challenged with WSSV, Negative control: Injection with buffer, not challenged with WSSV.
Presence of WSSV among dead and surviving individuals was tested using PCR at the end of the trial. DNA was extracted from the dead and surviving individuals. Pooled DNA samples of five individuals per tank were tested for the presence of WSSV. While all negative control samples (not challenged with WSSV) were free of virus, all positive control samples (not treated with dsRNA) tested positive for WSSV (Figure 2A and 2B). DNA from all RNAi treatment groups was PCR negative for the virus. This is a strong indication that treatment of shrimp with dsRNA prior to WSSV exposure inhibited viral replication and infection, resulting in effective protection against WSSV (Figure 2A and 2B).
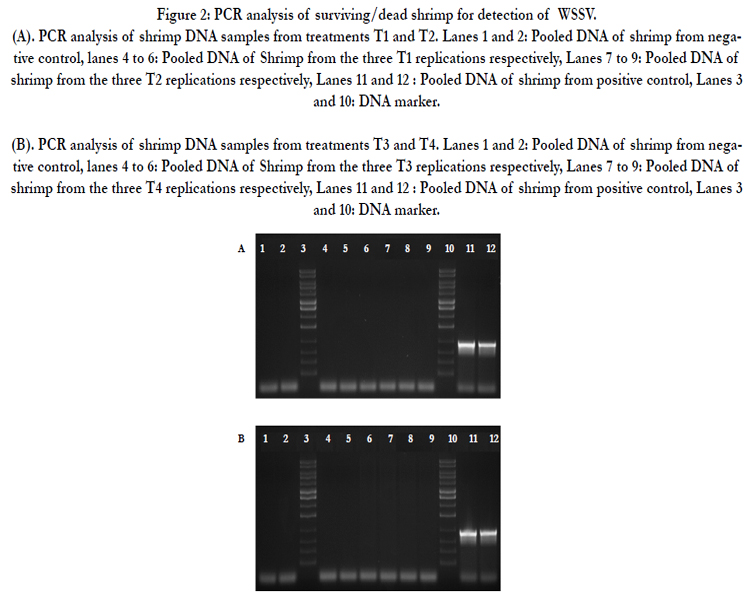
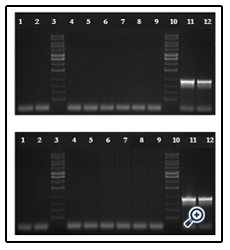
Figure 2: PCR analysis of surviving/dead shrimp for detection of WSSV.
(A). PCR analysis of shrimp DNA samples from treatments T1 and T2. Lanes 1 and 2: Pooled DNA of shrimp from negative control, lanes 4 to 6: Pooled DNA of Shrimp from the three T1 replications respectively, Lanes 7 to 9: Pooled DNA of shrimp from the three T2 replications respectively, Lanes 11 and 12 : Pooled DNA of shrimp from positive control, Lanes 3 and 10: DNA marker.
(B). PCR analysis of shrimp DNA samples from treatments T3 and T4. Lanes 1 and 2: Pooled DNA of shrimp from negative control, lanes 4 to 6: Pooled DNA of Shrimp from the three T3 replications respectively, Lanes 7 to 9: Pooled DNA of shrimp from the three T4 replications respectively, Lanes 11 and 12 : Pooled DNA of shrimp from positive control, Lanes 3 and 10: DNA marker.
Discussion
The key to development of an RNAi-based therapeutic (preferably oral) against WSSV depends on the ability of functional RNAi elements to provide effective protection to shrimp against WSSV for an extended duration. This would allow a shrimp farmer to apply the therapeutic once a week to protect the shrimp crop from WSSV infection. This is the first study that demonstrated that functional RNAi elements targeting WSSV genes delivered by injection remain active in shrimp for more than 14 days post-treatment and provide effective protection (more than 75% survival) to shrimp against WSSV for up to 28 days postdelivery of RNAi elements.The positive outcome reported in this study provides further impetus for development and refinement of RNAi-based therapeutics. Protection of up to 94% was observed in the present study when shrimp were challenged with WSSV 14 days post-injection of dsRNA targeting multiple WSSV genes. Only one previous study has evaluated the effectiveness of dsRNA targeting vp26 and vp28 on limiting WSSV infection 10 days and 20 days post treatment, reporting 20 to 37% and 0 to 13% survival when challenged with virus 10 days and 20 days after dsRNA treatment, respectively [17]. The increased protection (in terms of % survival as well as duration of protection) observed in the present study compared to the previous studies (Table 1) may have been due to the fact that four WSSV genes were targeted simultaneously in the present study, as opposed to targeting of a single WSSV genes [20, 25, 29, 17]. While two of the gene targets, vp28 and vp19 encode structural proteins, the other two gene targets rr2 and tk-tmk encode proteins involved in DNA replication. The simultaneous targeting of these two different sets of genes with broad functional characterisitics perhaps more fully inhibited viral replication which improved efficacy of the treatment, since WSSV was not detected among the surviving shrimp from these treatments.
The present study also sought to address another key question towards developing an RNAi-based therapeutic, which is whether it is best to provide a single large dosage or multiple small dosages of the RNAi elements. The results demonstrate that delivery of a single large dosage or multiple small dosages of dsRNA provide similar levels of protection. Since the RNAi elements provided to shrimp are expected to undergo degradation over time, the strategy of delivering small dosages of RNAi elements regularly appears to be more pragmatic. In addition, delivery of large dosages of RNAi elements is much more difficult to accomplish compared to periodic delivery of small dosages of dsRNA through transgenic bacteria, yeast or algae. This result is also encouraging since it is expected that oral delivery, the preferred route of delivery of this therapeutic, would only be capable of delivering small amounts of material over an extended period of time. This would allow a shrimp farmer to apply the RNAi-based therapeutic similarly to how they currently administer medicated feeds
Conclusion
In conclusion, the present study demonstrated that:
(i) injection delivery of RNAi elements targeting WSSV genes can provide protection to shrimp up to 28 days.
(ii) RNAi elements remain active in shrimp for up to 14 days post-delivery and,
(iii) multiple small dosages of RNAi elements are as effective as a single large dosage in providing effective protection. The practical implication of these results is that they provide valuable information for development of an RNAi-based therapeutic against WSSV.
References
- Aquatic Manual 2009 World organization for animal health (OIE). www.oie.int/
- Baum JA, Bogaert T, Clinton W, Heck GR, Feldmann P, et al. (2007) Control of coleopteran insect pests through RNA interference. Nature Biotechnol 25(11): 1322-1326.
- Chou HY, Huang CY, Wang CH, Chiang HC, Lo CF (1995) Pathogenicity of a baculovirus infection causing white spot syndrome in cultured penaeid shrimp in Taiwan. Dis Aqua Organ 23: 165–173.
- Davidson BL, McCray Jr PB (2011) Current prospects for RNA interference- based therapies. Nat Rev Genet 12(5): 329-340.
- Escobedo-Boniall CM (2011) Application of RNA interference (RNAi) against viral infections in shrimp: A review. J AntivirAntiretrovir S9: 2-12.
- Escobedo-Bonilla CM, Alday-Sanz V, Wille M, Sorgeloos P, Pensaert MB, et al. (2008) A review on the morphology, molecular characterization, morphogenesis and pathogenesis of white spot syndrome virus. J Fish Dis 31(1):1–18.
- Fire A, Xu SQ, Montgomery MK, Kostas SA, Driver SE, et al. (1998) Potent and specific genetic interference by double-stranded RNA in Caenorhabditiselegans. Nature 391(6669): 806–811.
- Hammond SM, Bernstein E, Beach D, Hannon GJ (2000) An RNA-directed nuclease mediates post-transcriptional gene silencing in Drosophila cells. Nature 404(6775): 293–296.
- Hammond S M, Boettcher S, Caudy AA, Kobayashi R, Hannon GJ (2001) Argonaute2, a link between genetic and biochemical analyses of RNAi. Science 293(5532): 1146–1150.
- Huang G, Allen R, Davis EL, Baum TJ, Hussey RS (2006) Engineering broad root-knot resistance in transgenic plants by RNAi silencing of a conserved and essential root-knot nematode parasitism gene. Proc Natl Acad Sci USA 103(39): 14302-14306.
- Jha RK, Xu ZR, Shen J, Bai SJ, Sun JY, et al. (2006) The efficacy of recombinant vaccines against white spot syndrome virus in Procambarusclarki. Immunol Lett 105(1): 68–76.
- Kumar A, Wang S, Ou R, Samarkandi M, Beerntsen BT, et al. (2013) Development of RNAi based microalgal larvicide to control mosquitoes. Malaria World Journal 4: 6.
- Lightner DV (1996) A handbook of pathology and diagnostic procedures for diseases of penaeid shrimp. World Aquaculture Society, Baton Rouge, LA.
- Lo CF, Ho CH, Peng SE, Chen CH, Hsu HC, et al. (1996) White spot syndrome baculovirus (WSBV) detected in cultured and captured shrimp, crabs and other arthropods. Dis Aqua Organ 27: 215-225.
- Martinez J, Patkaniowska A, Urlaub H, Luhrmann R, Tuschl T (2002) Single-stranded antisense siRNAs guide target RNA cleavage in RNAi. Cell 110(5): 563–574.
- Matzke MA, Birchler JA (2005) RNAi-mediated pathways in the nucleus. Nat Rev Genet 6(1): 24-35.
- Mejia-Ruiz CH, Vega-Pena S, Alvarez-Ruiz P, Escobedo-Bonilla CM (2011) Double-stranded RNA against white spot syndrome virus (WSSV) vp28 or vp26 reduced susceptibility of Litopenaeus vannamei to WSSV, and survivors exhibited decreased susceptibility in subsequent re-infections. J Invert Path 107(1): 65-68.
- NamikoshiA, Wu JL, Yamashita T, Nishizawa T, Nishioka T, et al. (2004) Vaccination trials with Penaeusjaponicus to induce resistance to white spot syndrome virus. Aquaculture 229(1): 25–35.
- Robalino J, Browdy CL, Prior S, Metz A, Parnell P, et al. (2004) Induction of antiviral immunity by double-stranded RNA in a marine invertebrate. J Virol 78(19): 10442–10448.
- RobalinoJ, Bartlett T, Shepard E, Prior S, Jaramillo G, et al. (2005) Doublestranded RNA induces sequence-specific antiviral silencing in addition to nonspecific immunity in a marine shrimp: Convergence of RNA interference and innate immunity in the invertebrate antiviral response. JVirol 79 (21): 13561–13571.
- Sarathi M, Simon MC, Venkatesan C, Sahul Hameed AS (2008) Oral administration of bacterially expressed VP28dsRNA to protect Penaeusmonodon from white spot syndrome virus. Marine Biotechnol(NY) 10 (3): 242-249.
- Sioud M (2015) RNA Interference: Mechanisms, technical challenges, and therapeutic opportunities. Methd Mol Biol 1218: 1-15.
- Van Hulten MCW, Witteveldt J, Peters S, Kloosterboer N, Tarchini R, et al (2001) The white spot syndrome virus DNA genome sequence. Virology 286(1): 7–22.
- Wang CH, Lo CF, Leu JH, Chou CM, Yeh PY, et al. (1995) Purification and genomic analysis of baculovirus associated with white spot syndrome (WSBV) of Penaeus monodon. Dis Aqua Organ 23(3): 239–242.
- Westenberg M, Heinhuis B, Zuidema D, Vlak JM (2005) siRNA injection induces sequence independent protection in Penaeus monodon against white spot syndrome virus (WSSV). Virus Res 114(1-2): 133–139.
- Witteveldt J, Cifuentes CC, Vlak JM, van Hulten MCW (2004) Protection of Penaeus monodon against white spot syndrome virus by oral vaccination. J Virol 78(4): 2057–2061.
- Wongteerasupaya C, Vickers JE, Sriurairatana S, Nash GL, Akarajamorn A, et al. (1995) A non-occluded, systemic baculovirus that occurs in cells of ectodermal and mesodermal origin and causes high mortality in the black tiger prawn Penaeus monodon. Dis Aqua Organ 21(1): 69–77.
- Wu Y, Lu L, Yang L, Weng S, Chan S, et al. (2007) Inhibition of white spot syndrome virus in Litopenaeus vannamei shrimp by sequence-specific siRNA. Aquaculture 271(1): 21-30.
- Xu J, Han F, Zhang X (2007) Silencing shrimp white spot syndrome virus (WSSV) genes by siRNA. Antiviral Research 73(2): 126–131
- Yadav BC, Veluthambi K, Subramaniam K (2006) Host-generated double stranded RNA induces RNAi in plant-parasitic nematodes and protects the host from infection. Mol Biochem Parasitol 148(2): 219-22.
- Yang F, He J, Lin X, Li Q, Pan D, et al. (2001) Complete genome sequence of the shrimp white spot bacilliform virus. J Virol 75(23): 11811–11820.





