Bacteriocin Producing Lactobacilli Strains as a Biological Strategy to Control Listerial Growth
García MJ1,2*, Ruiz F1,2, Asurmendi P1,2, Pascual L1, Barberis L1
1 Departamento de Microbiología e Inmunología, Universidad Nacional de Río Cuarto, Río Cuarto, Córdoba, Argentina.
2 Fellow of Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET).
*Corresponding Author
García, María José,
Departamento de Microbiología e Inmunología. Facultad de Ciencias
Exactas Físico-Químicas y Naturales. Universidad Nacional de Río Cuarto.
Ruta 36 Km. 601, Río Cuarto (5800), Argentina.
Tel: +54-0358-4676539
Fax: +54-0358-4676231
E-Mail: mjgarcia@exa.unrc.edu.ar
Recieved: August 19, 2015; Accepted: September 10, 2015; Published: September 12, 2015
Citation: García MJ, Ruiz F, Asurmendi P, Pascual L, Barberis L (2015) Bacteriocin Producing Lactobacilli Strains as a Biological Strategy toControl Listerial Growth. Int J Microbiol Adv Immunol. 03(2), 60-64. doi: dx.doi.org/10.19070/2329-9967-1500011
Copyright: García MJ© 2015. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
Listeria monocytogenes causes listeriosis in humans and animals, and is predominantly transmitted by ingestion of contaminated food. Its severity and high mortality rate make listeriosis a relevant foodborne disease. Currently, biological strategies using probiotic lactobacilli to prevent and control infectious diseases are being globally investigated. The biocontrol exerted by certain Lactobacillus spp. is mainly due to the production of a variety of antimicrobial substances. The aims of this work were (1) to investigate the presence of Listeria spp. in raw cow milk and (2) to determine the inhibitory activity of bacteriocin-like inhibitory substances (BLIS-es) produced by Lactobacillus fermentum L23 and Lactobacillus rhamnosus L60 on L. monocytogenes and other listerial species isolated from food and clinical samples. The presence of Listeria spp. in raw milk was evaluated in 814 samples from 238 cows. The listerial prevalence in cattle was 0.84% and the isolates were identified as L. innocua (LI1) and L. welshimeri (LW1). A total of 29 listerial strains were used as indicator microorganisms to evaluate the antimicrobial activity of BLIS-es L23 and L60. Bioactive metabolites produced by these lactobacilli strains were able to inhibit the listerial growth. This biological activity was mainly attributed to the BLIS-es L23 and L60 which, even at low concentration, were active on 100% of listerial strains. This study reveals a strong potential for the biotechnological use of these bacteriocin producing lactobacilli as a biostrategy against Listeria spp.
2.Introduction
3.Material and Methods
3.1 Isolation and identification of Listeria spp
3.2 Lactobacilli and culture media condition
3.3 Test of antimicrobial activity
3.4.Minimum inhibitory concentration of the bacteriocin like inhibitory substances on the listerial growth
3.5.Statistical Analysis
4.Results and Discussion
5.Conclusion
6.Acknowledgments
7.References
keywords
Antilisterial Activity; Bacteriocins; Lactobacillus spp.; Listeria monocytogenes; Raw Cow Milk.
Introduction
Listeria monocytogenes causes listeriosis in humans and animals, and is predominantly transmitted by ingestion of contaminated food. This microorganism can also be transmitted transplacentally from mother to child, during passage through the colonized birth canal, or after direct contact with infected animals. The major groups at risk of invasive listeriosis include pregnant women, neonates, elderly people and immunocompromised individuals. Clinical manifestations such as neonatal infections, abortions, bacteremia, meningitis and rhombencephalitis have been the most severe infections [1, 2]. Listeriosis is one of the most relevant foodborne diseases with an elevated socio-economical impact due to severity of invasive infections associated with a high mortality rate [3]. Listeria spp. are widely distributed in the environment and different foods, including dairy products. Recently, several outbreaks of listeriosis associated to consumption of dairy products have beenreported worldwide [4]. L. monocytogenes represents an important concern for food safety due to its ability to resist extreme conditions (low pH, high salt concentrations and refrigeration temperatures) which are commonly used as preservation procedures and thus may survive in food for long periods of time [2, 5]. Other listerial species, such as L. ivanovii, L. seeligeri, L. welshimeri and L. innocua, have been reported to cause infections in humans. Some of these species have been associated with bacteremia, acute meningitis, coagulation disorders, and multiple-organ dysfunction, leading to the death of the patients [6].
Although listeriosis is a notifiable disease in many developed countries, there has not been a reduction in its incidence [7, 8]. During the last years, the rate of L. monocytogenes infection has varied between 0.29 and 1.8 cases per 100,000 inhabitants [9, 10]. For this reason, other strategies of sanitary control against L. monocytogenes are being investigated. The application of lactobacilli exhibits a promising approach for the control of pathogenic microorganisms in both, the food industry as well as the biomedical field. The biocontrol exerted by Lactobacillus spp. is mainly due to the production of antimicrobial substances such as organic acids, hydrogen peroxide and bacteriocins [5, 11-13]. Bacteriocins are defined as proteinaceous antimicrobial substances, produced by bacteria that inhibit growth of related or unrelated bacterial species. Bacteriocin-like inhibitory substance (BLIS) is an acronym used to refer to those bacteriocins whose amino-acid sequences have not yet been elucidated [13]. Our research group has identified and characterized two lactobacilli strains, Lactobacillus fermentum L23 and Lactobacillus rhamnosus L60, which were selected for their bacteriocinogenic and probiotic properties. Furthermore, extensive investigations have demonstrated the ability of these strains to inhibit the growth of several pathogenic microorganisms [13-18]. Currently, researchers have become increasingly interested in the search for natural antimicrobial substances, such as bacteriocins, to develop biotechnological products for food industry applications as alternative to reduce chemical and thermal preservation methods [11, 19].
The aims of this work were (1) to investigate the presence of Listeria spp. in raw cow milk and (2) to determine the inhibitory activity of BLIS-es produced by Lactobacillus fermentum L23 and Lactobacillus rhamnosus L60 on L. monocytogenes and other listerial species isolated from food and clinical samples.
Material and Methods
Raw milk samples were collected between August 2012 and April 2014 from Villa María, Córdoba. 814 samples from different quarters of the mammary gland of 238 cows were analyzed for the presence of Listeria spp.
The isolation of Listeria spp. from milk samples was performed by means of the double enrichment method followed by a plate isolation procedure. The first selective enrichment of Listeria spp. was done in tryptic soy broth (TSB) (Britania, Argentina) added with ceftazidime (0.002% w/v) and trypaflavine neutral (0.25% w/v). Cultures were incubated for 48 h at 37°C. The enrichment broths were kept at 4°C for 48 h as a second selective enrichment. Each culture broth was seeded on Oxford agar plates (Britania, Argentina) and incubated at 37°C for 24-48 h. Typical colonies were identified by biochemical tests [20]. Other L. monocytogenes strains previously isolated from food and clinical samples were provided by the Bacteriology laboratory of Universidad Nacional de Río Cuarto, Argentina. They were seeded on tryptic soy agar (TSA) plates (Britania, Argentina) and incubated at 37°C for 24 h. Strains were stored at -80°C in TSB containing 30% (v/v) glycerol. All Listeria spp. from different origins were used as indicator microorganisms to evaluate the antimicrobial activity of the bioactive substances produced by L. fermentum L23 and L. rhamnosus L60.
Two human Lactobacillus strains, L. fermentum L23 and L. rhamnosus L60, were previously identified by standard biochemical tests, the API 50 CHL system (BioM`erieux, Inc., France) and 16S rRNA analysis. The 16S rRNA sequences of both lactobacilli were deposited in the GenBank under the accession numbers GQ 455406 and EF 495247 to L. fermentum L23 and L. rhamnosus L60, respectively [18]. Lactobacilli strains were grown in De Man Rogosa Sharpe (MRS) broth or agar (Britania, Argentina) at 37°C under a 5% CO2 atmosphere for 24 h. They were stored at -80°C in MRS broth containing 30% (v/v) glycerol.
Both Lactobacillus strains produce different antimicrobial compounds such as organic acids, bacteriocins and, in the case of L.rhamnosus L60, also hydrogen peroxide. The BLIS-es L23 and L60 were previously purified by at least three-step procedure developed for class II bacteriocins [14-16].
The antilisterial activity of L. fermentum L23 and L. rhamnosus L60 was tested by the streak diffusion method described by Asurmendi et al. [12]. On the other hand, the inhibitory activity of cell free supernatant (CFS) and treated or neutralized CFS (NCFS), containing the BLIS-es L23 or L60, was evaluated against Listeria spp. strains by well diffusion test, on agar plates [18]. L. fermentum L23 and L. rhamnosus L60 were cultured in MRS broth and incubated at 37°C under a 5% CO2 atmosphere for 20h. Then, the supernatants were removed by centrifugation (4,000xg at 4°C for 20 min). These fractions with biological activity were neutralized with NaOH 1 mol ml-1 to eliminate the inhibitory effects of the organic acids. TSA plates were seeded with a Listeria spp. culture (1.5 x 108 CFU ml-1) and wells were made into agar plates. 100 μl of CFS and NCFS from each Lactobacillus strain were added to different wells. Plates were incubated for 24 h at 37°C and inhibition halos were measured.
The minimum inhibitory concentration (MIC) values of each BLIS, L23 and L60, were evaluated using a modification of the well diffusion test on agar plates [16]. To obtain the BLIS L23, the CFS of L. fermentum L23 was neutralized with NaOH 1 mol ml-1. In the case of BLIS L60, the NCFS of L. rhamnosus L60 was also treated with 0.1 mg ml-1 peroxidase (Sigma) to eliminate the inhibitory effects attributed to the hydrogen peroxide. A suspension of Listeria spp. in TSB broth (1.5 x 108 CFU ml-1) was seeded on TSA plates. 100 μl of two fold serial dilutions of each BLIS were spotted on different agar wells and plates were incubated. MIC of both BLIS-es was defined as the reciprocal of the highest dilution which produced complete inhibition of the indicator growth, and was expressed as activity units per milliliter (AU ml-1) [21].
All tests were performed in triplicate, and mean ± SD were expressed. Differences in inhibitory activities between bacteriocin producing strains and their different bioactive supernatants were analyzed by ANOVA (P<0.05) using InfoStat Software. A P value of <0.05 was considered statistically significant.
Results and Discussion
In the present study, a total of 814 raw milk samples from 238 cows were examined for the presence of Listeria spp. Two samples, obtained from different cows, were positive for Listeria spp. The listerial prevalence percentage in cattle was 0.84%. According to the biochemical identification, the isolates were identified as L. innocua (LI1) and L. welshimeri (LW1). Our results agreed with a previous report by Aygun and Pehlivanlar [22], who found a low prevalence value in raw cow milk. In contrast, a significantly higher listerial prevalence has been reported in the US (23%) and Iran (22.5%) during the last years [4, 23]. The listerial species isolated in this work coincided with recent works reported by Jamali et al. [4] and Rahimi et al. [24] who recovered L. innocua and L. welshimeri among other listerial species.
Bacteria that belong to Listeria genus are indirect indicators of the potential presence of L. monocytogenes in food because all listerial species occur in similar environmental niches [25]. Since L. innocua and L. welshimeri have been reported to cause infections in humans, the presence of these Listeria spp. found in this work, would consequently imply that the consumption of raw or improperly pasteurized milk could represent a potential risk for human health. Moreover, the main causes of contamination for these bacteria in raw milk are associated with fecal and environmental sources. Such contaminations could occur during milking, storage or transport, or directly from infected animals in dairy farms [26]. Since the listerial species isolated in the present study are non-pathogenic for cows, we suggest that the presence of these microorganisms could be associated to environmental contaminations during milking practices, which are related with a poor hygienic quality of milk. In Argentina, the information about the prevalence of L. monocytogenes and other Listeria spp. in milk, is still extremely limited, bearing in mind, the detection of these microorganisms in milk is not regulated by the Código Alimentario Argentino [27]. To our knowledge, the only previous work carried out in our country to determine the presence of Listeria spp. in raw milk was reported by Laciar et al. [28]. These authors isolated L. monocytogenes, L. innocua and L. welshimeri. Considering that Argentina is an important producer of milk worldwide, the detection of listerial species, such as the one carried out in this work, shows that it is essential to search for these bacteria in milk in order to ensure its safety.
In this work, we report the in vitro antimicrobial activity of two probiotic and bacteriocinogenic strains, L. fermentum L23 and L. rhamnosus L60, on a total of 29 listerial strains (27 strains of L. monocytogenes isolated from food and clinical samples, and L. innocua LI1 and L. welshimeri LW1 recovered from raw cow milk). The streak diffusion method performed served as a preliminary technique for screening of listerial susceptibility to antimicrobial substances produced by these lactobacilli strains. The results showed that lactobacilli inhibited 100% of the tested Listeria spp. (data not shown). The inhibition zone produced by L. fermentum L23 showed a mean value of 20.64 ± 4.92 mm, whereas that of L. rhamnosus L60 was of 19.67 ± 4.94 mm. There was no statistical difference between these inhibition values (P<0.05). These results showed that both Lactobacillus strains had a strong biological activity to inhibit the listerial growth. Previous studies have demonstrated the antimicrobial power of different lactobacilli strains against certain species of Listeria spp. [29]. Nevertheless, in comparison with our work, L. fermentum L23 and L. rhamnosus L60 proved to have a wider range of antilisterial activity because they inhibited all Listeria spp. tested, independently from where they were originally isolated.
The antilisterial activity of the CFS and NCFS containing the BLIS-es, L23 or L60, from L. fermentum L23 or L. rhamnosus L60, respectively, was tested by the well diffusion method. The mean inhibition halos obtained on each listerial strain are shown in Table 1. Results showed that both CFS containing all the substances with antimicrobial activity inhibited all the tested Listeria spp. The mean inhibition halos produced by the CFS of L. fermentum L23 and L. rhamnosus L60 on Listeria spp. were 21.10 ± 1.94 and 21.08 ± 2.35, respectively. There was no statistical difference between these inhibition values (P<0.05). After treatment with NaOH 1 mol ml-1, both NCFS maintained a high antimicrobial activity on listerial growth. The mean inhibition halos produced by the NCFS of L. fermentum L23 and L. rhamnosus L60 on listerial growth were17.90 ± 1.74 mm and 17.73 ± 1.93 mm, respectively. Antimicrobial activities found with each NCFS against Listeria spp. growth did not show significant differences (P<0.05). The higher percentage of antilisterial activity remained in the NCFSs of both lactobacilli strains. In the case of L. fermentum L23 this biological activity was due to the BLIS L23 while for L. rhamnosus L60, the NCFS contains the BLIS L60 as well as hydrogen peroxide. Our research group have previously demonstrated that hydrogen peroxide produced by L. rhamnosus L60 had a weak antimicrobial activity on a wide variety of bacterial genera [13, 15, 21]. On this regards, it was assumed that the antimicrobial effect of both NCFSs was due to the BLIS-es L23 and L60, which were the metabolites responsible of the main antilisterial effect. Indeed, both bacteriocins inhibited the growth of 100% of Listeria spp. Only in the case of L. monocytogenes, similar results were found by Altuntas et al. [11], who demonstrated the susceptibility of those strains to the non-treated CFS containing a bacteriocin produced by Pediococcus spp. In a similar study, Vera Pingitore et al. [30] studied the antimicrobial activity of two NCFS containing different bacteriocins and found different levels of resistance among L. monocytogenes, L. innocua, L. welshimeri and L. seeligeri strains. Furthermore, Dortu et al. [31] reported a high sakacin G resistance level (40%) for L. monocytogenes strains. At this point, our findings demonstrate the relevant antilisterial activity of these two bacteriocins based on both, the large number of susceptible strains and the high sizes of inhibition halos produced by them.
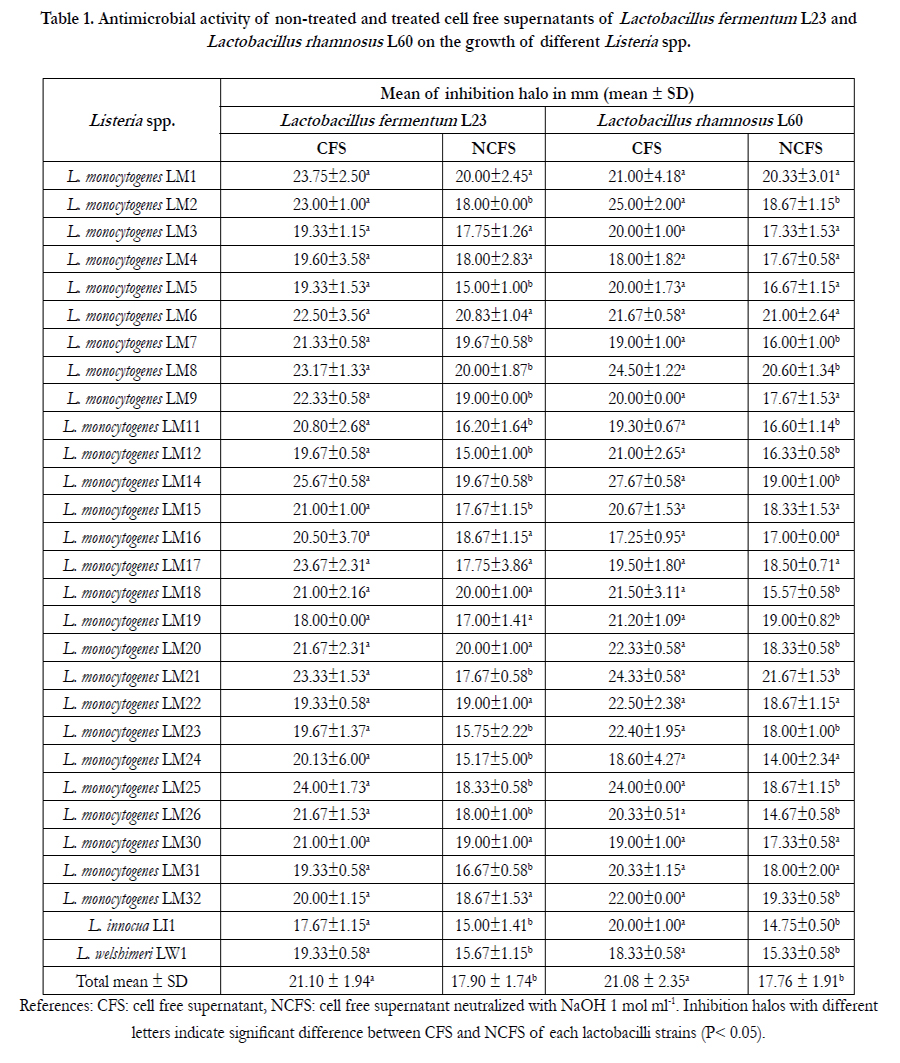

Table 1. Antimicrobial activity of non-treated and treated cell free supernatants of Lactobacillus fermentum L23 and Lactobacillus rhamnosus L60 on the growth of different Listeria spp.
The proportion of antimicrobial effect of the main biometabolites was estimated based on the sizes of inhibition halos produced by CFS and NCFS of each Lactobacillus strain. The average sizes of inhibition zones produced by CFS and NCFS of L. fermentum L23 strain were significantly different (P<0.05), due to the joint action of organic acid and the BLIS L23, in comparison with the inhibition found by the BLIS alone. Thus, the proportion of biological activity on Listeria spp. produced by BLIS L23 and organic acids were 85% and 15%, respectively. The same differences were observed between CFS and NCFS of L. rhamnosus L60. In this case, 84% of inhibition was mainly attributed to BLIS L60 and 16% to organic acids.
Table 2 shows the MIC values of BLIS-es L23 or L60 on all Listeria spp. MICs of both BLIS-es ranged between 40 and 160 AU ml-1 for all strains tested of Listeria spp. The BLIS L23 inhibited the growth of 19 strains of L. monocytogenes (70.37%) with a very low MIC value. In the case of the BLIS L60, 23 strains of L. monocytogenes (85.18%) were inhibited with MIC values ranged between 40-80 AU ml-1. On the other hand, both BLIS-es showed the same MIC value to inhibit the growth of L. innocua LI1 and L. welshimeri LW1. The low MIC values for each BLIS, in comparison with their maximum activity (640 AU ml-1), showed that even when they are highly diluted, they remained active against all Listeria spp. tested. These results differed from those by Kaur et al. [32], who evaluated pediocin 34 and enterocin FH99, whose MIC values were higher in comparison with the reported here for L23 and L60. These findings could serve as an biotechnological advantage for the development of novel products, potentially useful, to minimize the presence of Listeria spp. in food industry.
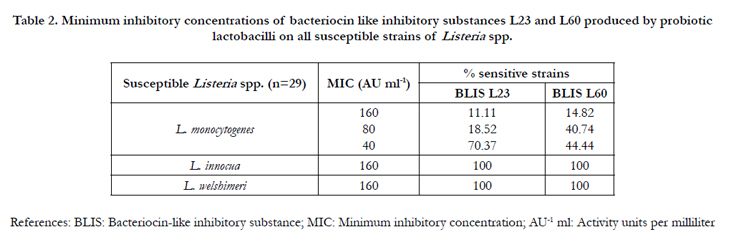
Table 2. Minimum inhibitory concentrations of bacteriocin like inhibitory substances L23 and L60 produced by probiotic lactobacilli on all susceptible strains of Listeria spp.
Conclusion
This study demonstrated that Listeria species contaminated raw cow milk in an important milking area of Argentina. In addition, L. fermentum L23 and L. rhamnosus L60 proved to have meaningful antimicrobial activity on all of the tested listerial strains. This antilisterial effect was mainly due to bacteriocins L23 and L60, which were still active at very low concentrations. These findings are promising as a biological strategy to prevent or reduce the risk of acquiring severe infections by the main pathogen, L. monocytogenes, through food transmission to human. Future studies shall be needed to evaluate the application of these bacteriocin producing strains in milk and/or dairy food.
Acknowledgments
This work was supported by the Secretaría de Ciencia y Técnica, Universidad Nacional de Río Cuarto, Córdoba, Argentina. M. J. García has a doctoral fellowship from CONICET (Consejo Nacional de Investigaciones Científicas y Técnicas), Argentina. Dr. F Ruiz and Dr. P. Asurmendi have a posdoctoral fellowship from CONICET.
References
- Allerberger F, Wagner M (2010) Listeriosis: a resurgent foodborne infection. Clin Microbiol Infect 16(1): 16-23.
- Sip A, Wieckowicz M, Olejnik-Schmidt A, Grajek W (2012) Anti-Listeria activity of lactic acid bacteria isolated from golka, a regional cheese produced in Poland. Food Control 26(1): 117-124.
- Muñoz AI, Vargas M, Otero L, Díaz G, Guzmán V (2011) Presencia de Listeria monocytogenes en alimentos listos para el consumo, procedentes de plazas de mercado y delicatessen de supermercados de cadena, Bogotá, D.C, 2002-2008. Biomédica 31(3): 428-439.
- Jamali H, Radmehr B, Thong KL (2013) Prevalence, characterisation, and antimicrobial resistance of Listeria species and Listeria monocytogenes isolates from raw milk in farm bulk tanks. Food Control 34(1): 121-125.
- Cosentino S, Fadda ME, Deplano M, Melis R, Pomata R, et al. (2012) Antilisterial activity of nisin-like bacteriocin-producing Lactococcus lactis subsp. lactis isolated from traditional sardinian dairy products. J Biomed Biotechnol 2012: 1-8.
- Perrin M, Bemer M, Delamare C (2003) Fatal case of Listeria innocua bacteremia. J Clin Microbiol 41(11): 5308-5309.
- Vrbova L, Johnson K, Whitfield Y, Middleton D (2012) A descriptive study of reportable gastrointestinal illnesses in Ontario, Canada, from 2007 to 2009. BMC Public Health 12: 970.
- Centers for Disease Control and Prevention (CDC) (2013a) Incidence and trends of infection with pathogens transmitted commonly through foodfoodborne diseases active surveillance network, 10 U.S. sites, 1996-2012. MMWR 62(15): 283-287.
- Jensen AK, Ethelberg S, Smith B, Nielsen EM, Larsson J, et al. (2010) Substantial increase in listeriosis, Denmark 2009. Euro Surveill 15(12): 19522.
- Centers for Disease Control and Prevention (CDC) (2013b) Vital Signs: Listeria illnesses, deaths, and outbreaks-United States, 2009-2011. MMWR 62(22): 448-452.
- Altuntas EG, Kocan D, Cosansu S, Ayhan K, Juneja VK, et al. (2012) Antibiotic and bacteriocin sensitivity of Listeria monocytogenes strains isolated from different foods. Food Nutr Sci 3(3): 363-368.
- Asurmendi P, García MJ, Pascual L, Barberis L (2015) Biocontrol of Listeria monocytogenes by lactic acid bacteria isolated from brewer´s grains used as feedstuff in Argentina. J Stored Prod Res 61: 27-31.
- Ruiz FO, Pascual L, Giordano W, Barberis L (2015) Bacteriocins and other bioactive substances of probiotic lactobacilli as biological weapons against Neisseria gonorrhoeae. Pathog Dis 73(3): 1-10.
- Pascual LM, Daniele MB, Giordano W, Pájaro MC, Barberis IL (2008a) Purification and partial characterization of novel bacteriocin L23 produced by Lactobacillus fermentum L23. Curr Microbiol 56(4): 397-402.
- Pascual LM, Daniele MB, Ruiz F, Giordano W, Pájaro C, et al. (2008b) Lactobacillus rhamnosus L60, a potential probiotic isolated from human vagina. J Gen Appl Microbiol 54(3): 141-148.
- Ruiz FO, Gerbaldo G, Asurmendi P, Pascual LM, Giordano W, et al. (2009) Antimicrobial activity, inhibition of urogenital pathogens, and synergistic interactions between Lactobacillus strains. Curr Microbiol 59(5): 497-501.
- Daniele M, Ruiz F, Pascual L, Barberis L (2011) Ureaplasma urealyticum and Mycoplasma hominis sensitivity to bacteriocins produced by two lactobacilli strains. Curr Microbiol 63(4): 360-365.
- Ruiz FO, Gerbaldo G, García MJ, Giordano W, Pascual L, et al. (2012) Synergistic effect between two bacteriocin-like inhibitory substances produced by lactobacilli strains with inhibitory activity for Streptococcus agalactiae. Curr Microbiol 64(4): 349-356.
- Khanian SI, Mojgani N, Ahmedi MK (2014) Characterization of partially purified bacteriocin like substance (BLIS) produced by probiotic Lactobacillus strains. Int J Enteric Pathog 2(2): e17426.
- Vos P, Garrity G, Jones D, Krieg NR, Ludwin W, et al. (2009) Bergey´s Manual of Systematic Bacteriology. The Firmicutes. (2nd edtn), Springer, New York. 3: 244-257.
- Ruiz F (2013) Efecto terapéutico de las bacteriocinas producidas por las cepas de Lactobacillus fermentum y Lactobacillus rhamnosus sobre microorganismos causales de infecciones genitales. Doctoral Thesis. UNRC. Río Cuarto, Córdoba, Argentina.
- Aygun O, Pehlivanlar S (2006) Listeria spp. in the raw milk and dairy products in Antakya, Turkey. Food Control 17(8): 676-679.
- Latorre AA, Van Kessel JAS, Karns JS, Zurakowski MJ, Pradhan AK, et al. (2009) Molecular ecology of Listeria monocytogenes: evidence for a reservoir in milking equipment on a dairy farm. Appl Environ Microbiol 75(5): 1315-1323.
- Rahimi E, Momtaz H, Sharifzadeh A, Behzadnia A, Ashtari MS, et al. (2012) Prevalence and antimicrobial resistance of Listeria species isolated from traditional dairy products in Charar Mahal & Bakhtiary, Iran. Bulgarian Journal of Veterinary Medicine 15(2): 115-122.
- Suh SH, Jaykus LA (2013) Nucleic acid aptamers for capture and detection of Listeria spp. J Biotechnol 167(4): 454-461.
- Sharma D, Sharma PK, Saharan BS, Malik A (2012) Isolation, identification and antibiotic susceptibility profiling of antimicrobial resistant Listeria monocytogenes from dairy milk. Int J Microb Resour Technol 1(1): 1-4.
- Código Alimentario Argentino (CAA) Chapter VIII: Alimentos lácteos. Updated 2014. Publishing ANMAT (Administración Nacional de Medicamentos, Alimentos y Tecnología Médica) website: www.anmat.gov.ar/alimentos/normativas_alimentos_caa.asp.
- Laciar AL, Vaca L, de Centorbi P (1999) Listeria spp. en alimentos de origen animal. Rev Argent Microbiol 31(1): 25-30.
- Chen H, Hoover DG (2003) Bacteriocins and their food applications. Comprehensive Reviews in Food Science and Food Safety 2(3): 82-100.
- Vera Pingitore E, Todorov SD, Sesma F, Franco BD (2012) Application of bacteriocinogenic Enterococcus mundtii CRL35 and Enterococcus faecium ST88Ch in the control of Listeria monocytogenes in fresh Minas cheese. Food Microbiol 32(1): 38-47.
- Dortu C, Huch M, Holzapfel W, Franz CMAP, Thonart P (2008) Antilisteria activity of bacteriocin-producing Lactobacillus curvatus CWBI-B28 and Lactobacillus sakei CWBI-B1365 on raw beef and poultry meat. Lett Appl Microbiol 47(6): 581-586.
- Kaur G, Singh TP, Malik RK (2013) Antibacterial efficacy of Nisin, Pediocin 34 and Enterocin FH99 against Listeria monocytogenes and cross resistance of its bacteriocin resistant variants to common food preservatives. Braz JMicrobiol 44(1): 63-71.





