Role of Adipocytes in Obesity and Diabetes: Its Potential Therapeutic Prospects
Saxena M*, Modi DR
Department of Biotechnology, Babasaheb Bhimrao Ambedkar University (A Central University), Lucknow, India.
*Corresponding Author
Dr. Madhukar Saxena,
Department of Biotechnology, Babasaheb Bhimrao Ambedkar University (A Central University),
Vidya Vihar, Rai Bareilly Road, Lucknow-226025, India.
E-mail: madhukar@bbau.ac.in; madhukarbio@gmail.com
Received: July 15, 2016; Accepted: September 23, 2016; Published: September 26, 2016
Citation: Saxena M, Modi DR (2016) Role of Adipocytes in Obesity and Diabetes: Its Potential Therapeutic Prospects. Int J Diabetol Vasc Dis Res,. 4(4), 166-170. doi: dx.doi.org/10.19070/2328-353X-1600035
Copyright: Saxena M © 2016. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
Obesity is the epidemic of the 21st century while type 2 diabetes mellitus (T2DM) is one of the most prevalent chronic diseases worldwide and increasing global burdens of disease day by day. Obesity and overweight increase the risk of several serious chronic diseases, such as T2DM, hypercholesterolemia, hypertriglyceridemia, hypertension, arthritis, asthma, cardiovascular disease, stroke and certain types of cancer. Adipocytes have a capability of a metabolic control and endocrine organs that have both good as well as bad effects. This is more clearly define the role adipocytes play in obesity and how inflammatory mediators act as signaling molecules in this process. Adiponectin is a serum protein produced and secreted exclusively by adipose tissues. In humans, plasma adiponectin concentrations fall with increasing obesity and reduced adiponectin concentrations correlate with insulin resistance and hyperinsulinemia. Adipocytes undergoes molecular and cellular alterations affecting systemic metabolism when an individual have enlarged adipocytes and become obese. Mesenchymal stem cells possess immunomodulatory activities (acting on both innate and acquired immunity effectors) which should result in a reduction of the immunogenicity of transplanted cells which lead to the destruction of beta cells. Currently stem cell of various source are under investigation for transplantable insulin-producing cells. In conclusion, it is now apparent that adipocytes are not simply a storage reservoir of fat but are active endocrine organs that play multiple roles in the body. Increases in inflammation and various cytokines from the adipose tissue, activate monocytes enhancing insulin resistance which increasing the risk for diabetes. Finally, the intracellular mechanisms by which these factors affect energy intake, utilization and metabolism are being better understood and we are developing therapies that manipulate these pathways.
2.Introduction
3.Role of Adipokines In Obesity and Diabetes
4.Molecular and Cellular Aspects of Obesity and Diabetess
5.Role of Mesenchymal Stem Cells and Its Therapeutic Potential
6.Conclusion
7.Acknowledgements
8.References
Key Words
Obesity; Type 2 Diabetes Mellitus; Adipocytes; Mesenchymal Stem Cells; Therapeutics.
Introduction
Obesity is the epidemic of the 21st century while type 2 diabetes mellitus (T2DM) is one of the most prevalent chronic diseases worldwide, contributing significantly to the global burden of disease [1]. Impaired insulin secretion, increased hepatic glucose production, and decreased peripheral glucose utilization are the core defects responsible for the development and progression of T2DM (Figure 1). In developing countries, the prevalence of obesity continues to rise and obesity is occurring at younger ages. The World Health Organization Statistics Report estimates that globally one in six adults in obese and 2.8 million individuals die each year due to overweight or obesity [2]. Obesity and overweight increase the risk of several serious chronic diseases, such as T2DM, cardiovascular disease, hypertension and stroke, hypercholesterolemia, hypertriglyceridemia, arthritis, asthma and certain forms of cancer [3, 4]. Present consideration on the role of fat itself by adipocytes (Fat storage cells) in the development of obesity and its consequences was considered to be a submissive. However, adipocytes are a critical component of metabolic control and endocrine organs that have both good and bad effects. This is more clearly define the role adipocytes play in health and in obesity and how inflammatory mediators act as signaling molecules in this process. Moreover, at a molecular level, we are beginning to understand how such variables as hormonal control, exercise, food intake and genetic variation interact which result in a given phenotype. Chronic inflammation is implicated in the pathogenesis of T2DM and in the development of cardio vascular disease (CVD) and other diabetic complications including diabetic retinopathy [5]. Inflammatory cytokines and adipocytokines secreted by adipocytes are involved in the regulation of glucose metabolism and insulin resistance and also in other inflammatory processes linked to an increased T2DM and CVD risk [6] such as high levels of C-reactive protein (CRP) are related to risk of future CVD in people with T2DM [7]. The inflammatory nature of T2DM is partly mediated through increased adiposity [8], reduced levels of adiponectin [9], an anti-inflammatory cytokine with anti-atherogenic properties with hepatic CRP secretion suggested to increase in response to an adiposity related increase in another inflammatory cytokine, interleukin-6 (IL-6).
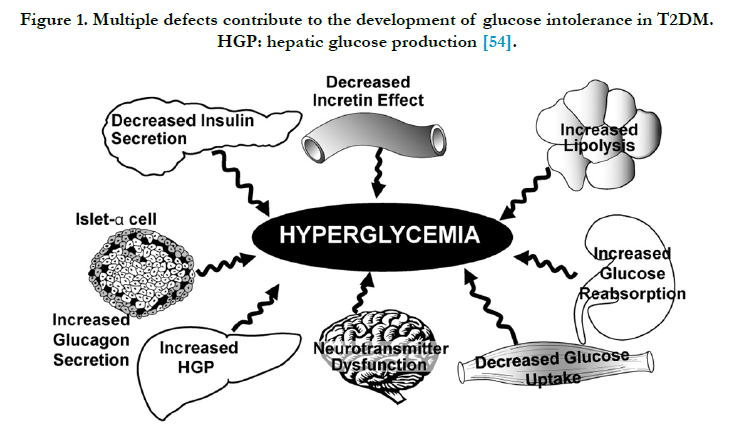
Figure 1. Multiple defects contribute to the development of glucose intolerance in T2DM. HGP: hepatic glucose production [54].
Role of Adipokines In Obesity and Diabetes
The present knowledge attributes that the role of adipose tissue is energy storage in the form of fatty acids. The metabolic role of adipocytes is however, complex involving inflammatory processes viz. leptin, resistin, visfatin, etc. The effects of specific proteins may be autocrine or paracrine, or the site of action may be distant from adipose tissue. Resistin is an adipose tissue-specific factor which is reported to induce insulin resistance, linking diabetes to obesity [10]. Adiponectin is a serum protein produced and secreted exclusively by adipose tissues. Adiponectin also known as adipocyte complement related protein of 30 KDa (Acrp30), is a hormone of adipocyte origin that is involved in the homeostatic control of circulating glucose and lipid level [11,12]. Adiponectin is a 147 amino acid protein that is similar in sequence and structure to the C1q complement factor. This hormone enhances insulin sensitivity in muscle and liver and increases free fatty acid (FFA) oxidation in several tissues, including muscle fibers [13-15]. It also decreases serum FFA, glucose and triacylglycerol concentrations. In humans, plasma adiponectin concentrations fall with increasing obesity and this effect is greater in men than in women [16]. Reduced adiponectin concentrations correlate with insulin resistance and hyperinsulinemia [17,18]. In addition, several polymorphisms of the adiponectin gene have been identified that are associated with reduced plasma adiponectin concentration and that increase the risk of T2DM, insulin resistance or metabolic syndrome [11].
Interestingly, adiponectin appears to be implicated in the development of atherosclerosis. Adiponectin concentrations are reduced in patients with CVD and adiponectin inhibits tumor necrosis factor α (TNF- α) induced expression of adhesion molecules and the transformation of macrophages to foam cells, both of which are key components of atherogenesis [19]. Leptin, the first adipocyte hormone identified, influences food intake through a direct effect on the hypothalamus [20]. Unlike insulin and leptin, adiponectin levels in plasma remain constant throughout the day and are not acutely affected by food intake. In humans and rodents, plasma leptin concentrations are highly correlated with BMI [21]. Leptin is the protein product of ob gene in mice and is involved in appetite control. Lack of gene coding for leptin (ob/ob mice) in mice are very obese and diabetic [22]. Mice and rats with a genetic mutation affecting the leptin receptor in the hypothalamus exhibit a similar phenotype to ob/ ob mice [23]. However, as animals and humans become obese, the role of leptin in regulating body weight becomes more complex. There are certainly rare cases where mutations affecting the genes coding for either leptin or its receptor have been found in families with a high prevalence of morbid obesity [24-26]. However, in most obese individuals, leptin concentrations are already high because of the increased amount of leptin-secreting adipose tissue. It appears that with increasing leptin concentrations, the hormone induces target cells to become resistant to its actions. Humans with a rare disorder called lipoatrophic diabetes have little or no fat mass, reduced serum adipokines such as leptin and very elevated serum triacylglycerol concentrations. In fact, triacylglycerol concentrations tend to be in the thousands, so high that some individuals require regular plasmapheresis to reduce serum triacylglycerol. These elevated lipid concentrations lead to an enlarged fatty liver, which can lead to severe liver disease and some individuals die secondary to liver complications. Clearly, both adiponectin and leptin are important hormones with both central and peripheral effects on metabolism and energy balance. Resistin is another hormone secreted by adipocytes that acts on skeletal muscle myocytes, hepatocytes and adipocytes themselves, where it is suggested to reduce insulin sensitivity leading to T2DM[9,11]. Visfatin is considered a new member of the adipokine family. It is highly expressed in visceral fat and whose level correlates with obesity [11]. The action and identity of diabetes susceptibility genes represent an important area for consideration of disease mechanism.
Molecular and Cellular Aspects of Obesity and Diabetes
Adipose tissue undergoes molecular and cellular alterations affecting systemic metabolism when an individual have enlarged adipocytes and become obese. First, fasting whole-body FFA and glycerol release from adipocytes is increased in obese women compared with lean women which probably promotes insulin resistance [27]. Increased FFAs are well known to promote insulin resistance in tissues like in muscle [28]. One of the common cause for the increased release of FFA is secondary to alterations in expression of perilipins. Perilipins are phosphoproteins found in adipocytes on the surface of triacylglycerol droplets that act as gatekeepers, preventing lipases from hydrolyzing triacylglycerol to facilitate the release of FFAs [29]. Perilipins is deficit in obese individuals even if their fat cells are larger, hence their increased basal rate of lipolysis [30]. However, several proinflammatory factors are produced in adipocytes with increasing obesity. Adipocytes of obese shows higher expression of proinflammatory proteins, including TNF-α, interleukin-6 (IL-6), monocyte chemotactic protein 1, inducible nitric oxide synthase, transforming growth factor β1, procoagulant proteins such as plasminogen activator inhibitor type 1, tissue factor and factor VII in comparison with lean individuals [31-34]. The chronological appearance of these inflammatory molecules before the development of insulin resistance and other complications of obesity, strongly suggests adipose tissue inflammation as an important protagonist in the development of obesity and T2DM and its associated complications. Inflammation is thought to contribute to the development of the sequelae of obesity. Certain cytokines are thought to reduce adiponectin expression [35]. TNF-α may increase systemic insulin resistance by promoting the release of fatty acids from adipose tissue into the bloodstream to act on tissues such as muscle and liver. Thus, adipose tissue TNF-α can act locally in adipose tissue, which ultimately promotes insulin resistance in peripheral tissues. However, IL-6 expression is also increased in obese adipose tissue. Plasma concentrations of IL-6 increase with obesity, unlike those of TNF-α, which acts in an autocrine and paracrine fashion [36]. IL-6 increases lipolysis and fat oxidation in humans and plasma IL-6 concentrations correlate with insulin resistance. Elevated IL-6 concentration is a predictor for development of T2DM [37, 38]. Some endocrinologists previously posited that obesity results from an increased endogenous production of the glucocorticoid hormone cortisol. It is well known that either endogenous overproduction of cortisol or exogenous administration of corticosteroids results in weight gain, with an increase in visceral fat deposition compared with peripheral fat (central obesity). Hypercortisolinemia, additionally, results in hyperphagia, central obesity, high concentrations of VLDL, insulin resistance and predisposition to diabetes. Clearly, several inflammatory mediators are implicated in the development of obesity and the mechanisms responsible for the development of the chronic diseases associated with obesity including diabetes and metabolic syndrome.
Role of Mesenchymal Stem Cells and Its Therapeutic Potential
Mesenchymal stem cells (MSCs) are uniquely capable of crossing germinative layers borders and are viewed as promising cells for regenerative medicine approaches in several diseases. MSCs are obtainable in high numbers via ex vivo culture and can be differentiated towards insulin producing cells (IPCs) [3]. Moreover, recent reports evidenced that MSCs possess immunomodulatory activities (acting on both innate and acquired immunity effectors) which should result in a reduction of the immunogenicity of transplanted cells, thus limiting rejection. Moreover it has been proposed that MSCs administration should be used to attenuate the autoimmune processes which lead to the destruction of beta cells [39]. Historically, the focus on MSCs has been defining their ability to differentiate into cells of the mesenchymal lineage and their subsequent potential therapeutic use [40]. However, a lot of pathophysiological abnormalities targeted by currently available antidiabetic medications are reviewed (Figure 2). In recent years there has been a shift in focus towards understanding the therapeutic potential of the secretions from MSCs [41]. The lack of intracellular adiponectin accumulation suggests that metformin is able to up-regulate adiponectin release by modulating posttranslational events [42]. In contrast to this the effect of anti-diabetic drugs, it could be hypothesized that the non-adipose components of subcutaneous adipocytes participate in the regulation of metformin effects on adiponectin production and secretion. This hypothesis agrees with the inability of metformin therapy to modify in vivo adiponectin content and secretion in a model of isolated adipocytes of obese diabetic patients. It could therefore hypothesized that metformin merely influences tissue availability of adiponectin through a decrease in the local inflammatory state [41]. Pioglitazone increased the gene expression of several factors involved in mitochondrial biogenesis. Pioglitazone increased the expression of genes required for fatty acid oxidation. The expression of homolog implicated in the regulation of fatty acid oxidation tended to be higher after pioglitazone treatment [43].
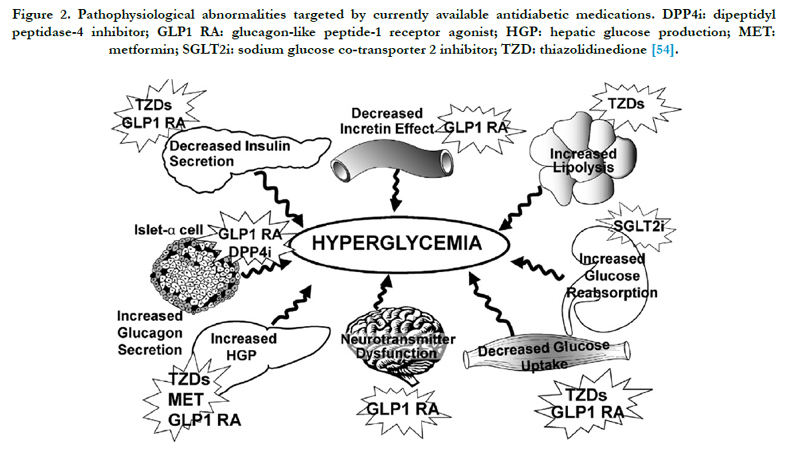
Figure 2. Pathophysiological abnormalities targeted by currently available antidiabetic medications. DPP4i: dipeptidyl peptidase-4 inhibitor; GLP1 RA: glucagon-like peptide-1 receptor agonist; HGP: hepatic glucose production; MET: metformin; SGLT2i: sodium glucose co-transporter 2 inhibitor; TZD: thiazolidinedione [54].
Consequently, as part of normal tissue function, there is significant cross-talk between the cells, which occurs though direct cell-cell contact or surface molecular receptors and secreted factors. Indeed, co-culture experiments have demonstrated complex reciprocal signaling, between adipose-derived cells [44]. In previous studies with SVF and adipocyte cells are interacting in a manner that enhances their paracrine signaling by inducing the cytokine production. The inflammatory secretion data support that hypothesis and demonstrate that analyzing individual cell populations will provide an incomplete understanding of the cytokine activity of adipose tissue [4]. The in vitro culturing of ADSCs resulted in the secretion of significantly higher concentrations of cytokines (IFN-γ, IL-7, IL-10, IL-12, IL-13 and VEGF) when compared to the co-culture dataset. Consequently, the increased secretion of some cytokines by the isolated adherent ADSCs, when compared to co-culture samples, may be due to the increase in cell numbers during culture and the lack of immune cells and adipocytes regulating their secretions [45-47]. Consequently the secretion profiles produced by these isolated and mixed cell populations are of interest in order to delineate functional differences and guide potential therapeutic use [48]. However, sufficient amounts of HUMSCs in rat livers can secrete cytokines, reduce the activation of hepatic stellate cells, enhance liver cell repair and effectively cure liver fibrosis [49].
Elevated serum proinflammatory cytokine levels have been attributed, in part, to the increased number of resident macrophages within adipose tissue of obese individuals. One such report suggests that undifferentiated and/or adipocyte differentiated ASCs within intact adipose tissues also may contribute to the elevated proinflammatory cytokines levels found in obese individuals [50]. Moreover, in one of our study results demonstrated the new information on cholesterol, triglycerides and lipoproteins release from adipocytes indicates that the post obese complication in in vivo will increase the disease associated with lipid metabolism [38]. The effect of anti-diabetic drugs on adipose tissue represents an additional mechanism through which this may induce clinical benefits [51]. Furthermore, obesity is characterized by elevated serum levels of pro-inflammatory and chemotactic cytokines.
Traditionally the management of diabetes is achieved by conventional anti-diabetic therapies including insulin treatment and self blood glucose monitoring can significantly prevent the development and retard the progression of chronic associated complications. Replacement of a patient’s islets (β-cells) is the only treatment of type 1 diabetes (T1DM) that achieves an insulin-independent, constant normoglycaemic state and avoids hypoglycaemic episodes [52]. However, islet transplantation has been hampered by the worldwide shortage of transplant-ready islets, immune rejection and recurrent attacks against islets by the underlying autoimmunity [53]. Currently stem cell of various source are under investigation for transplantable insulinproducing cells. Human mesenchymal stem cells derived from Wharton’s Jelly may serve as a promising alternative cell source of transplantable islet-like clusters and its potential to become an excellent candidate in β-cell replacement therapy of diabetes [54].
Conclusion
In conclusion, it is now apparent that adipocytes are not simply a storage reservoir of fat but are active endocrine organs that play multiple roles in the body. Their metabolic role changes as they enlarge with increasing obesity. This increased understanding of the role of the adipocyte and its associated adipokines, such as leptin and adiponectin, is allowing us to dissect the all too prevalent metabolic syndrome and perhaps affect its course for the better. Diabetes is one of the major causes of global morbidity and mortality. Over the last two decades an avalanche of information has emerged on the sensory apparatus and signaling cas-cades that mobilize innate immunity in response to diabetes. Increases in inflammation and various cytokines from the adipose tissue, activate monocytes enhancing insulin resistance which increasing the risk for diabetes. Although current diabetes management focuses on lowering blood glucose, the goal of therapy should be to delay disease progression and eventual treatment failure. Recent innovative treatment approaches target the multiple pathophysiological defects present in T2DM. We are also beginning to understand the interplay of inflammation and obesity, although our knowledge remains incomplete. Finally, the intracellular mechanisms by which these factors affect energy intake, utilization and metabolism are being better understood and we are developing therapies that manipulate these pathways.
Acknowledgements
Madhukar is thankful to National Post Doctoral Fellowship from Department of Science and Technology Science and Engineering Research Board, New Delhi (PDF/2015/000007). The central equipment facility of Department of Biotechnology, BBA University is duly acknowledged.
References
- Chen L, Magliano DJ, Zimmet PZ (2011) The worldwide epidemiology of type 2 diabetes mellitus present and future perspectives. Nat Rev Endocrinol. 8(4): 228-36.
- World Health Organization (WHO). World Health Statistics 2. 2012. Geneva.
- Anzalone R, Lo Iacono M, Loria T, et al., (2011) Wharton's jelly mesen chymal stem cells as candidates for beta cells regeneration: extending the differentiative and immunomodulatory benefits of adult mesenchymal stem cells for the treatment of type 1 diabetes. Stem Cell Rev 7(2): 342-63.
- Biancone L, Bruno S, Deregibus MC, et al., (2012) Therapeutic potential of mesenchymal stem cell-derived microvesicles. Nephrol Dial Transplant. 27(8): 3037–3042.
- Williams M, Nadler J (2007) Inflammatory mechanisms of diabetic complications. Curr Diab Rep. 7(3): 242-8.
- Pearson TA, Mensah GA, Alexander RW, et al., (2003) Markers of inflammation and cardiovascular disease: application to clinical and public health practice: a statement for healthcare professionals from the Centers for Disease Control and Prevention and the American Heart Association. Circulation.107(3): 499-511.
- Yu HI, Sheu WHH, Song YM, et al., (2004) C-reactive protein and risk factors for peripheral vascular disease in subjects with type 2 diabetes mellitus. Diabet Med. 21(4): 336-41.
- Kriketos AD, Greenfield JR, Peake PW, et al., (2004) Inflammation, insulin resistance, and adiposity: a study of first-degree relatives of type 2 diabetic subjects. Diabetes Care. 27(8): 2033-40.
- Silha J, Krsek M, Skrha J, et al., (2003) Plasma resistin, adiponectin and leptin levels in lean and obese subjects: correlations with insulin resistance. Eur J Endocrinol. 149(4): 331-5.
- Trayhurn P, Beattie JH (2001) Physiological role of adipose tissue: white adipose tissue as an endocrine and secretory organ. Proc Nutr Soc. 60(3): 329-39.
- Saxena M, Srivastava N, Banerjee M (2012) Genetic association of adiponectin gene polymorphisms (+45T/G and +10211T/G) with type 2 diabetes in North Indians. Diabetes Metab Syndr. 6: 65-69.
- Scherer PE, Williams S, Fogliano M, et al., (1995) A novel serum protein similar to C1q, produced exclusively in adipocytes. J Biol Chem. 270(45): 26746-9.
- Fruebis J, Tsao TS, Javorschi S, et al., (2001) Proteolytic cleavage product of 30-kDa adipocyte complement-related protein increases fatty acid oxidation in muscle and causes weight loss in mice. Proc Natl Acad Sci. 98: 2005–10.
- Yamauchi T, Kamon J, Waki H, et al., (2001) The fat-derived hormone adiponectin reverses insulin resistance associated with both lipoatrophy and obesity. Nat Med. 7: 941-6
- Scherer PE, Williams S, Fogliano M, et al., (1995) A novel serum protein similar to C1q, produced exclusively in adipocytes. J Biol Chem. 270: 26746-9.
- Arita Y, Kihara S, Ouchi N, et al., (1999) Paradoxical decrease of an adiposespecific protein, adiponectin, in obesity. Biochem Biophys Res Commun. 257(1): 79-83.
- Hotta K, Funahashi T, Arita Y, et al., (2000) Plasma concentrations of a novel, adipose-specific protein, adiponectin, in type 2 diabetic patients. Arterioscler Thromb Vasc Biol. 20(6): 1595-9.
- Hotta K, Funahashi T, Bodkin NL, et al., (2001) Circulating concentrations of the adipocyte protein adiponectin are decreased in parallel with reduced insulin sensitivity during the progression to type 2 diabetes in rhesus monkeys.Diabetes. 50: 1126-33.
- Ouchi N, Kihara S, Arita Y, et al., (2001) Adipocyte-derived plasma protein, adiponectin, suppresses lipid accumulation and class A scavenger receptor expression in human monocyte-derived macrophages. Circulation 103:1057-63.
- Lee GH, Proenca R, Montez JM, et al., (1996) Abnormal splicing of the leptin receptor in diabetic mice. Nature 379: 632-5.
- Maffei M, Halaas J, Ravussin E, et al., (1995) Leptin levels in human and rodent: measurement of plasma leptin and ob RNA in obese and weightreduced subjects. Nat Med. 1: 1155-61.
- Pelleymounter MA, Cullen MJ, Baker MB, et al., (1995) Effects of the obese gene product on body weight regulation in ob/ob mice. Science. 269: 540-3.
- Chua SC Jr, Chung WK, Wu-Peng XS, et al., (1996) Phenotypes of mouse diabetes and rat fatty due to mutations in the OB (leptin) receptor. Science. 271: 994-6.
- Montague CT, Farooqi IS, Whitehead JP, et al., (1997) Congenital leptin deficiency is associated with severe early-onset obesity in humans. Nature. 387: 903-8.
- Farooqi IS, Keogh JM, Kamath S, et al., (2001) Partial leptin deficiency and human adiposity. Nature. 414: 34-5.
- Clement K, Vaisse C, Lahlou N, Carbol S, Pelloux V, et al., (1998) A mutation in the human leptin receptor gene causes obesity and pituitary dysfunction. Nature. 392: 398-401.
- Horowitz JF, Coppack SW, Paramore D, et al., (1999) Effect of short-term fasting on lipid kinetics in lean and obese women. Am J Physiol. 276: E278- 84.
- Shulman GI (2000) Cellular mechanisms of insulin resistance. J Clin Invest. 106(2): 171-6.
- Zhang HH, Souza SC, Muliro KV, et al., (2003) Lipase-selective functional domains of perilipin A differentially regulate constitutive and protein kinase A-stimulated lipolysis. J Biol Chem. 278: 51535-42.
- Wang Y, Sullivan S, Trujillo M, et al., (2003) Perilipin expression in human adipose tissues: effects of severe obesity, gender, and depot. Obes Res. 11: 930-6.
- Perreault M, Marette A (2001) Targeted disruption of inducible nitric oxide synthase protects against obesity-linked insulin resistance in muscle. Nat Med. 7: 1138-43.
- Weyer C, Yudkin JS, Stehouwer CD, Pratley RE, et al., (2002) Humoral markers of inflammation and endothelial dysfunction in relation to adiposity and in vivo insulin action in Pima Indians. Atherosclerosis. 161: 233-42.
- Sartipy P, Loskutoff DJ (2003) Monocyte chemoattractant protein 1 in obesity and insulin resistance. Proc Natl Acad Sci. 100(12): 7265-70.
- Samad F, Pandey M, Loskutoff DJ (1998) Tissue factor gene expression in the adipose tissues of obese mice. Proc Natl Acad Sci. 95(13): 7591-6.
- Bruun JM, Lihn AS, Verdich C, Pedersen SB, Toubro S, et al., (2003) Regulation of adiponectin by adipose tissue-derived cytokines: in vivo and in vitro investigations in humans. Am J Physiol Endocrinol Metab. 285: E527-33.
- Mohamed-Ali V, Goodrick S, Rawesh A, Miles JM,Yudkin Js, et al., (1997) Subcutaneous adipose tissue releases interleukin-6, but not tumor necrosis factor-alpha, in vivo. J Clin Endocrinol Metab. 82(12): 4196-200.
- Pradhan AD, Manson JE, Rifai N, Buring JE, Ridker PM, et al., (2001) C-reactive protein, interleukin 6, and risk of developing type 2 diabetes mellitus. JAMA. 286(3): 327-34.
- Saxena M, Modi DR, Singh S, Prakash A (2015) Effect of glucose feed on cholesterol, triglycerides and lipoprotein concentrations in preadipocytes and adipocytes: an in-vitro study. Eur J Mol Biol Biochem. 2(3): 118-122.
- English K, French A, Wood KJ (2010) Mesenchymal Stromal Cells: Facilitators of Successful Transplantation? Cell Stem Cell. 7(4): 431-42.
- Blaber SP, Webster RA, Hill CJ, Breen JE, Kuah D, et al., (2012) Analysis of in vitro secretion profiles from adipose-derived cell populations. J Translational Med. 10: 172-88.
- Phillips SA, Kung J, Ciaraldi TP, Choe C, Mudaliar S, et al., (2009) Selective regulation of cellular and secreted multimeric adiponectin by antidiabetic therapies in humans. Am J Physiol Endocrinol Metab. 297(3): E767-73.
- Perrini S, Laviola L, Cignarelli A, Melchiorre M, De Stefano F, et al., (2008) Fat depotrelated differences in gene expression, adiponectin secretion, and insulin action and signalling in human adipocytes differentiated in vitrofrom precursor stromal cells. Diabetologia. 51(1): 155-64.
- Bogacka I, Xie H, Bray GA, Smith SR (2005) Pioglitazone induces mitochondrial biogenesis in human subcutaneous adipose tissue in vivo. Diabetes. 54(5): 1392-9.
- Nishimura S, Manabe I, Nagasaki M, Eto K, Yamashita H, et al., (2009) CD8+ effector T cells contribute to macrophage recruitment and adipose tissue inflammation in obesity. Nat Med. 15: 914-20.
- Bunnell BA, Flaat M, Gagliardi C, Patel B, Ripoll C (2008) Adipose-derived stem cells: isolation, expansion and differentiation. Methods. 45: 115-20.
- Rosen ED and MacDougald OA (2006) Adipocyte differentiation from the inside out. Nat Rev Mol Cell Biol. 7: 885-96.
- Tayal V and Kalra BS (2008) Cytokines and anti-cytokines as therapeutics– an update. Eur J Pharmacol. 579: 1-12.
- Tsai PC, Fu TW, Chen YM, et al., (2009) The Therapeutic Potential of Human Umbilical Mesenchymal Stem Cells from Wharton’s Jelly in the Treatment of Rat Liver Fibrosis. Liver Transpl. 15(5): 484-95.
- Kilroy GE, Foster SJ, Wu X, Ruiz J, Sherwood S, et al., (2007) Cytokine Profile of Human Adipose-Derived Stem Cells: Expression of Angiogenic, Hematopoietic, and Pro-Inflammatory Factors. J cellular physiol. 212: 702-9.
- Bretzel RG, Browatzki CC, Schultz A, et al., (1993) Clinical islet transplantation in diabetes mellitus-report of the Islet Transplant Registry and the Giessen Center experience. Diabet Stoffwechsel. 2: 378-90.
- Saxena M, Modi DR, Singh S, Prakash A (2015) The effect of anti-diabetic drugs on the inflammatory cytokines in human in vitro subcutaneous adipocytes. Eur J Pharm Med Res. 2(3): 796-812.
- Shapiro AM, Lakey JR, Ryan EA, Korbutt GS, Toth E, et al., (2000) Islet transplantation in seven patients with type 1 diabetes mellitus using a glucocorticoid- free immunosuppressive regimen. N Engl J Med. 343(4): 230-8.
- Chao KC, Chao KF, Fu YS , Liu SH (2008) Islet like clusters derived from mesenchymal stem cells in Wharton's Jelly of the human umbilical cord for transplantation to control type 1 diabetes. Plos one. 3(1): e1451.
- DeFronzo RA, Triplitt CL, Abdul-Ghani M, Cersosimo E (2014) Novel Agents for the treatment of Type 2 Diabetes. Diabetes Spectrum. 27(2): 100-12.





