Blueberry Tea Enhances Insulin Sensitivity by Augmenting Insulin- Mediated Metabolic and Microvascular Responses in Skeletal Muscle of High Fat Fed Rats
Dino Premilovac1, Katherine M. Roberts-Thomson1, Huei L.H. Ng1, Eloise A. Bradley1, Stephen M. Richards1,Stephen Rattigan1, Michelle A. Keske1*.
Menzies Research Institute Tasmania, University of Tasmania, Hobart, Australia.
*Corresponding Author
Michelle A. Keske
Menzies Research Institute Tasmania,
University of Tasmania, Hobart, Australia.
Tel: (03) 62 26 2669; Fax: (03) 62 26 7704
E-mail: Michelle.Keske@utas.edu.au
Article Type: Research Article
Received: November 21, 2013; Accepted: December 16, 2013; Published: December 18, 2013
Citation: Keske MA et al,. (2013) Blueberry Tea Enhances Insulin Sensitivity by Augmenting Insulin- Mediated Metabolic and Microvascular Responses in Skeletal Muscle of High Fat Fed Rats. Int J Diabetol Vasc Dis Res. 1(5), 29-36. doi: dx.doi.org/10.19070/2328-353X-130006
Copyright: Keske MA© 2013 This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
Background: The aim of the current study was to determine whether a unique blueberry tea (BT) ameliorates insulin resistance by improving metabolic and vascular actions of insulin in skeletal muscle.
Methods: Male Sprague Dawley rats were fed normal (4.8% fat wt/wt, ND) or high (22.6% fat wt/wt, HFD) fat diets for 4 weeks. A second group of HFD rats was provided BT (4.0% wt/vol) in the drinking water during the final 2 weeks. Animals were subjected to an intraperitoneal glucose tolerance test (1g glucose/kg IPGTT) or euglycaemic hyperinsulinaemic clamp (10mU/min/kg x 2hr).
Results: HFD rats displayed significantly (p< 0.05) higher plasma glucose levels at 15 and 30 mins following the IPGTT compared to ND, and this increase was completely abolished by BT treatment. Glucose infusion rate, muscle glucose uptake, and microvascular perfusion in muscle were significantly (p< 0.05) impaired during clamps in HFD and all markedly improved (p< 0.05) with BT treatment. Insulin-mediated changes in femoral artery blood flow were unaffected by HFD or BT treatment.
Conclusions: We conclude that BT treatment ameliorates glucose intolerance and insulin resistance by restoring both metabolic and microvascular insulin sensitivity in high fat-fed rats. Therefore, BT consumption may have therapeutic implications for insulin resistance and type 2 diabetes.
2.Introduction
3.Methods
3.1 Animal Care
3.1 Surgical Preparation
4 Results
4.1 Physical and biochemical characteristics of rats
5 Discussion
6.Conclusions
7.Competing Interests
8.Authors’ Contributions
9.Acknowledgements
10.References
Key Words
Insulin Sensitivity; Glucose Tolerance; High Fat Diet; Blueberry Tea; Microvascular Perfusion; Muscle Glucose Uptake.
Introduction
Insulin resistance plays a key role in the pathogenesis of type-2 diabetes (T2D). While the root causes of insulin resistance are multifactorial, two of the major contributors are increased dietary fat and a sedentary lifestyle [1-4]. A defining feature of insulin resistance is impaired glucose disposal by the skeletal muscle, which is normally responsible for ~80% of insulin-mediated glucose uptake in the post-prandial state [5].
In addition to the classical metabolic actions of insulin to stimulate glucose uptake in skeletal muscle, insulin also has important vascular actions [6-13]. Our research implicates vascular dysfunction in skeletal muscle as one major cause of muscle insulin resistance. Insulin stimulates both total blood flow to skeletal muscle [11-13] and increases microvascular perfusion of the myocytes [6-10]. However insulin’s actions on microvascular perfusion and total muscle blood flow are separate events and insulin-mediated glucose uptake is altered by changes in microvascular perfusion and not total muscle blood flow [7,14].
We have demonstrated that insulin infusion or the ingestion of a mixed meal stimulate microvascular blood flow in skeletal muscle in both experimental animals [6-10] and humans [15-17]. Insulin stimulates microvascular recruitment to facilitate delivery of glucose and insulin to the myocyte to enhance glucose disposal [6-10, 18]. Insulin resistant rats [3] and obese humans [15, 16] also display marked reductions in insulin-mediated microvascular recruitment and insulin-mediated muscle glucose uptake suggesting that the loss of muscle perfusion may contribute to the insulin resistance. Recently, we reported that high fat-induced insulin resistance can originate from impaired microvascular insulin responses and that this microvascular defect precedes the development of myocyte insulin resistance [14].
Current treatments for T2D are limited, have unwanted side effects, and lose effectiveness over time. Finding novel therapeutic treatments that act by improving microvascular and metabolic insulin responses in skeletal muscle may aid in the prevention of insulin resistance and T2D or potentially enhance the action of current treatments. There is a growing public interest in the use of complementary and alternative approaches for treating insulin resistance and T2D. Blueberries and blueberries leaves have been reported to have anti-diabetic actions [19-21] and have been used as a traditional medicine for glycaemic control [22, 23]. Here we report on novel findings demonstrating that a unique blueberry tea blend (BT) restores metabolic and microvascular insulin sensitivity in muscle of the high fat-fed rat model.
Male Sprague-Dawley rats (4 weeks of age) were obtained from the University of Tasmania Central Animal Facility. On arrival, the rats were divided into two groups and provided either a normal (4.8% fat wt/wt; ND, Specialty Feeds, Glen Forest, WA, Australia) or high (22.6% fat wt/wt; HFD, Specialty Feeds) fat diet ad libitum for 4 weeks. The macronutrients in these diets are shown in Table 1. A second group of HFD rats were supplemented with blueberry tea (HFD+BT; 4% wt/vol) during the final two weeks of HFD feeding. The blueberry tea, Blueberry Boost™, was a kind donation from Dromana Blueberries (Oyster Cove, Tasmania, Australia) which contains a propriety blend of dried blueberries, blueberry leaves, spearmint leaves, raspberry leaves and cinnamon. BT was brewed at the rate of 4g of loose leaves per 100ml of boiling water, allowed to infuse for 10 minutes, then strained through a 0.75mm mesh to remove particulate matter and cooled. Animals in the HFD+BT group were provided with both normal drinking water and BT in separate drinking bottles ad libitum. The tea was replaced every two days. All animals were housed at 21±2°C with a 12h-12h light-dark cycle. All experimental procedures were approved by the University of Tasmania Animal Ethics Committee and performed in accordance with the Australian Code of Practice for the Care and Use of Animals for Scientific Purposes – 2013, 8th Edition.
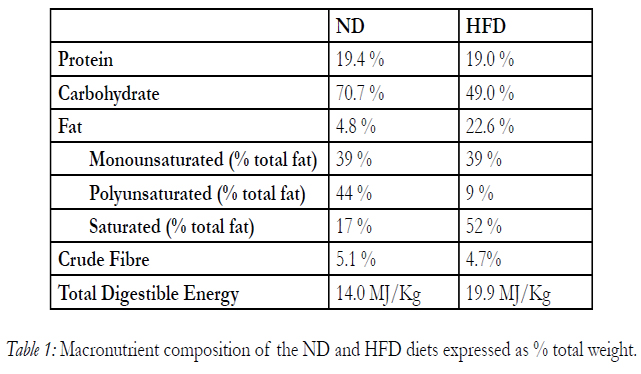

Table 1: Macronutrient composition of the ND and HFD diets expressed as % total weight.
Rats were anaesthetised with an intraperitoneal injection of aqueous pentobarbital sodium (50mg/kg body wt.) and surgery performed as previously described [6- 10]. Animals were maintained under anaesthesia for the duration of the experiment by an intravenous infusion of pentobarbital sodium (0.6 mg/min/kg body wt.), and mean arterial blood pressure (MAP) and heart rate continuously recorded. Following surgical preparation, 1 hr was allowed for the animal’s cardiovascular parameters to stabilise. Subsequently, animals were then subjected to either an intraperitoneal glucose tolerance test (protocol 1), or a euglycaemic hyperinsulinaemic clamp (protocol 2).
Animals were subjected to a 90 min glucose tolerance test (1g glucose/kg i.p.). Arterial blood was sampled from the carotid artery at baseline (0 min) and at 15,30, 45, 60 and 90 min following the glucose injection. Blood samples were centrifuged and plasma glucose levels were assessed using a glucose analyser (YSI 2300,Yellow Springs Instruments, OH, USA).
Following an overnight fast a 2 hr infusion of insulin (10mU/min/kg; Humulin, Eli Lilly, West Ryde, NSW, Australia) was initiated through the jugular vein. A 30% glucose solution (wt/vol) was infused at a variable rate to maintain fasting blood glucose concentrations over the course of the experiment. Arterial blood glucose levels were assessed from the carotid artery every 10-15 min using a glucose analyser (YSI 2300) and the glucose infusion rate (GIR) was adjusted accordingly.
Fasting and clamp plasma insulin concentrations were determined by ELISA (Mercodia AB, Uppsala, Sweden). Fasting plasma non esterified fatty acid (NEFA) levels were determined using an enzymatic assay kit (Wako Pure Chemical Industries, Osaka, Japan).
Muscle-specific glucose uptake was assessed by uptake of 2-deoxy-D-[1-14C]glucose (2DG; 0.1mCi/mL; American Radiolabeled Chemicals, Inc., MO, USA) as described previously [3, 14]. Briefly, 45 min prior to the conclusion of the experiment an i.v. bolus of 2DG was given (20μCi). Arterial plasma samples (25μL) were collected 5, 10, 15, 30 and 45 min after the 2DG bolus to assess plasma specific activity of 2DG. At the end of the experiment the calf muscle group (gastrocnemius, plantaris, soleus) was excised, immediately freeze clamped and stored at -80°C until analysed. The frozen muscle was ground under liquid nitrogen and ~100mg was homogenised with 1.5mL of distilled water. The homogenate was centrifuged and 1mL of supernatant was separated for free and phosphorylated 2DG using an anion exchange column (AG-1X8; Bio-Rad Laboratories, CA, USA). Biodegradable counting scintillant (Amersham, Arlington Heights, IL, USA) was added to each sample and radioactivity was measured using a scintillation counter (Tri-Carb 2800TR, Perkin Elmer, IL, USA). From this measurement and the specific activity of 2DG in plasma, the rate of muscle glucose uptake (R’g) was calculated as previously described [24].
Femoral artery blood flow (FBF) was continuously measured by an ultrasonic flow probe positioned around the femoral artery (Transonic, Ithica, NY, USA). Microvascular perfusion in muscle was assessed from metabolism of exogenously infused 1-methyl xanthine (1-MX; Sigma Aldrich, St. Louis, MO, USA) as previously published [3,6,8,14,25-29]. Briefly, a bolus of allopurinol (10μmol/kg; Sigma Aldrich) was given 5 min prior to 1-MX infusion to partially inhibit the activity of xanthine oxidase and ensure a constant saturating arterial level of 1-MX. 1-MX (0.4mg/ min/kg) was infused intravenously during the final 60min of the experiment. At the end of the experiment 100μL of arterial plasma was added to 20μL of perchloric acid (2M) to precipitate proteins. Hindleg venous plasma, obtained from the femoral vein using a 29G needle, was collected and 100μL was mixed with 20μL of perchloric acid (2M). The arterial and venous samples were centrifuged and the supernatant assessed for 1-MX and oxypurinol concentrations using reverse-phase high-performance liquid chromatography as previously described [6]. The metabolism of 1-MX was calculated from the plasma arterio-venous 1-MX difference multiplied by FBF and expressed as nmol/min.
Data are presented as the means ± SEM and statistics were performed using SigmaStat (Systat Software, San Jose, CA, USA). Comparisons between groups were made using One-way ANOVA. Comparison of timeseries measurements in each group was performed by two-way repeated measures ANOVA. When a significant difference of p< 0.05 was detected, pairwise comparisons by Student-Newman-Keuls test was used to assess treatment differences.
At the conclusion of the dietary intervention, there were no differences in body weight (Figure 1A) or energy intake from food (Table 2) between ND, HFD or HFD+BT groups. Water intake was the same between ND and HFD groups. However the HFD+BT group consumed more fluids as they were given both water and BT ad libitum (Table 2). At the end of the dietary intervention, there were no differences in plasma glucose concentrations between ND, HFD and HFD+BT rats (Figure 2A and B). As expected, HFD rats had significantly (p< 0.05) elevated epididymal fat pad weights (Figure 1B), fasting plasma insulin (Figure 3A), and fasting plasma NEFA (Figure 3B) concentrations when compared to ND. These outcomes were unaffected by BT treatment, however there was a trend (p=0.07) for fasting plasma NEFA concentrations to be lower in the HFD+BT when compared to HFD.
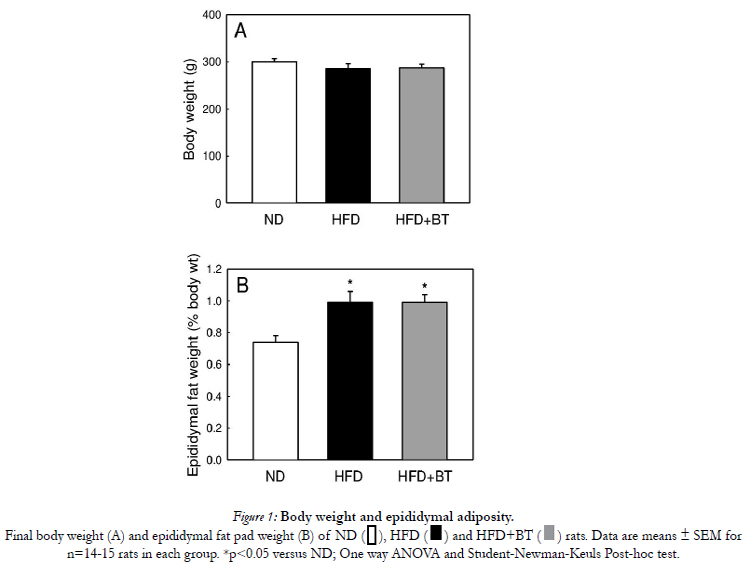
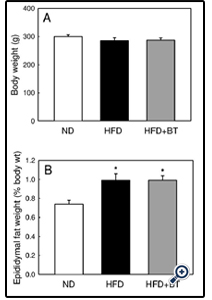
Figure 1: Body weight and epididymal adiposity.
Final body weight (A) and epididymal fat pad weight (B) of ND ,HFD and HFD+BT rats. Data are means ± SEM for n=14-15 rats in each group. *p < 0.05 versus ND; One way ANOVA and Student-Newman-Keuls Post-hoc test.
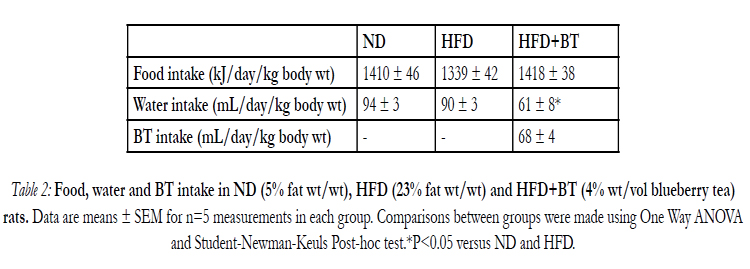
Table 2: Food, water and BT intake in ND (4.8% fat wt/wt), HFD (22.6% fat wt/wt) and HFD+BT (4% wt/vol blueberry tea) rats. Data are means ± SEM for n=5 measurements in each group. Comparisons between groups were made using One Way ANOVA and Student-Newman-Keuls Post-hoc test.*P < 0.05 versus ND and HFD.
Non-fasting (A) and fasting (B) plasma glucose levels in ND, HFD and HFD+BT rats. Data are means ± SEM for n=7-9 rats in each group. Data not significant. One way ANOVA .
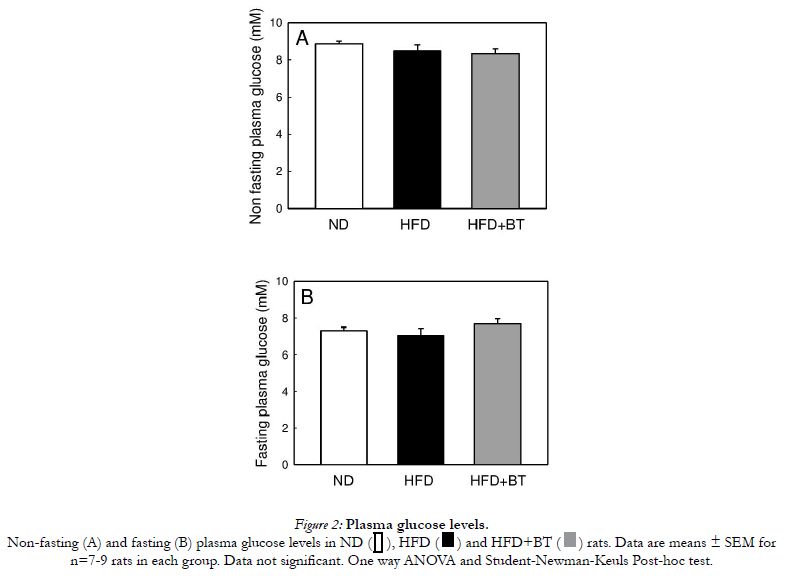
Figure 2: Plasma glucose levels.
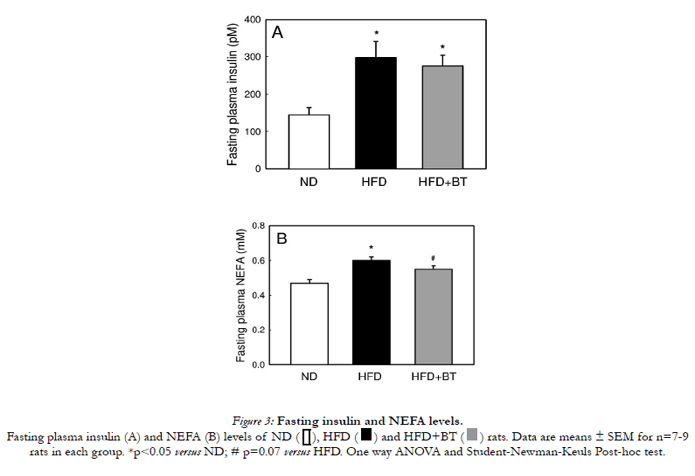
Figure 3: Fasting insulin and NEFA levels.
Fasting plasma insulin (A) and NEFA (B) levels of ND, HFD and HFD+BT rats. Data are means ± SEM for n=7-9 rats in each group. *p < 0.05 versus ND; # p=0.07 versus HFD. One way ANOVA and Student-Newman-Keuls Post-hoc test.
Figure 4 shows the time course of changes in plasma glucose following the glucose tolerance test. ND, HFD and HFD+BT groups had similar basal (non-fasting) plasma glucose levels. HFD had significantly higher plasma glucose at 15 and 30 min following the glucose challenge. Interestingly, the HFD+BT group had markedly (p< 0.05) reduced plasma glucose levels at 15 and 30 min when compared to HFD. However, there were no statistical differences in plasma glucose levels between groups at 0, 45, 60 or 90 min.
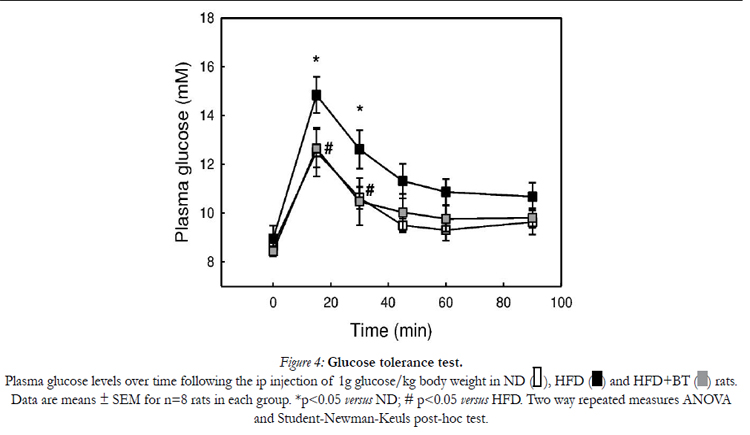
Figure 4: Glucose tolerance test.
Plasma glucose levels over time following the ip injection of 1g glucose/kg body weight in ND, HFD and HFD+BT rats. Data are means ± SEM for n=8 rats in each group. *p< 0.05 versus ND; # p< 0.05 versus HFD. Two way repeated measures ANOVA and Student-Newman-Keuls post-hoc test.
Figure 5A shows the GIR required to maintain glycaemia during insulin infusion for ND, HFD and HFD+BT groups. As expected, the HFD group had significantly lower (p< 0.05) GIR when compared to the ND group confirming that the HFD rats were insulin resistant. Importantly, the HFD+BT group had significantly (p< 0.05) elevated GIR when compared to HFD, indicating improved whole body insulin sensitivity. Insulin-mediated uptake of radiolabeled glucose in skeletal muscle (R’g, Figure 5B) was significantly (p< 0.05) impaired in HFD rats (18.6 ± 1.2 vs. 28.9 ± 2.9 μg/g/min, p < 0.05). However, insulin-stimulated R’g in HFD rats was markedly improved with BT treatment (24.4 ± 0.9 μg/g/min, p< 0.05).
Insulin increased FBF in all groups to a similar extent in ND, HFD and HFD+BT groups (p= NS, Figure 6A). Femoral artery vascular resistance (MAP divided by FBF) in response to insulin was also similar between groups (107 ± 15 vs. 120 ± 21 vs. 109 ± 15 mmHg.min/ml, ND, HFD and HFD+BT respectively, p= NS). Hindlimb 1-MX metabolism, an indicator of microvascular perfusion, was significantly (p< 0.05)impaired in HFD when compared to ND during insulin infusion (4.87± 0.63 vs. 8.43 ± 1.13 nmole/min; p< 0.05, Figure 6B). BT treatment reversed the impairment in insulin-mediated 1-MX metabolism induced by HFD (7.87 ± 0.63 nmole/min).
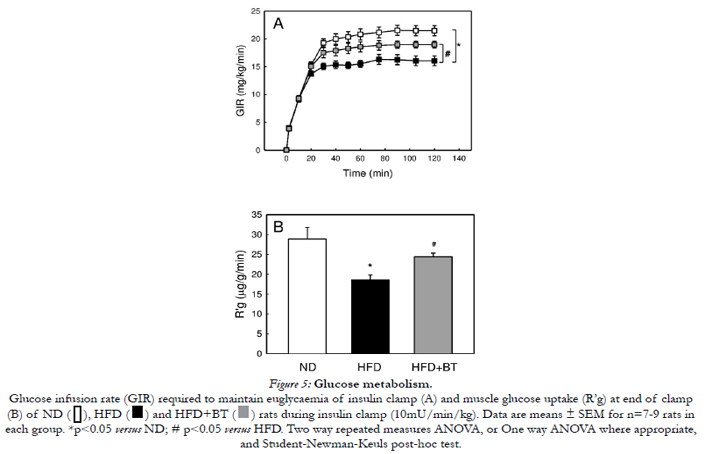
Figure 5: Glucose metabolism.
Glucose infusion rate (GIR) required to maintain euglycaemia of insulin clamp (A) and muscle glucose uptake (R’g) at end of clamp (B) of ND, HFD and HFD+BT rats during insulin clamp (10mU/min/kg). Data are means ± SEM for n=7-9 rats in each group. *p< 0.05 versus ND; # p< 0.05 versus HFD. Two way repeated measures ANOVA, or One way ANOVA where appropriate,and Student-Newman-Keuls post-hoc test.
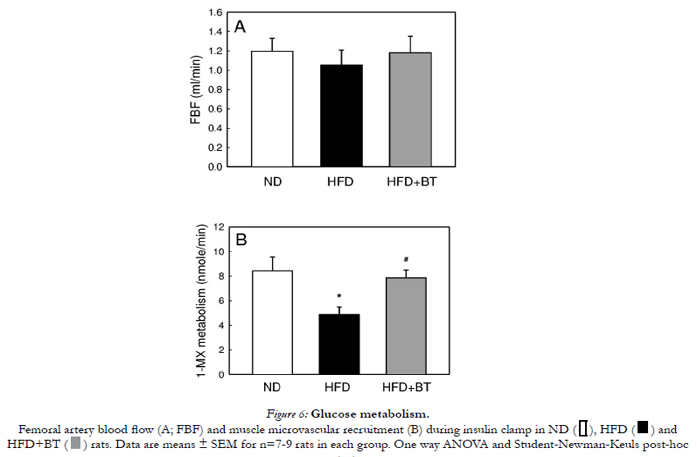
Figure 6: Glucose metabolism.
Femoral artery blood flow (A; FBF) and muscle microvascular recruitment (B) during insulin clamp in ND, HFD and HFD+BT rats. Data are means ± SEM for n=7-9 rats in each group. One way ANOVA and Student-Newman-Keuls post-hoc test.
*p<0.05 Versus ND; #p<0.05 Versus HFD
Discussion
The main findings from this study were that BT treatment ameliorated HFD-induced glucose intolerance and insulin resistance. BT treatment improved insulinstimulated glucose uptake and microvascular perfusion in skeletal muscle. These effects occurred without any changes in body weight, epididymal pad weight, fasting clinical chemistries or femoral artery blood flow. These data indicate that BT is an insulin sensitiser in skeletal muscle and acts by improving both the metabolic and microvascular actions of insulin.
Blueberry BoostTM is a proprietary blend of dried blueberries (37% wt/wt), blueberry leaves, raspberry leaves, spearmint leaves and cinnamon. Blueberries [19-21], blueberry leaves [20] and cinnamon [30-32] have each been reported to improve insulin sensitivity or insulin action; however there are no reports of raspberry leaves or spearmint on these measures. High fat fed mice supplemented with freeze-dried blueberry powder (4% wt/wt provided in the food x 8 weeks) prevented diet-induced obesity and improved whole body insulin sensitivity in response to an insulin tolerance test [19]. Blueberry fruit and blueberry leaf extracts have been shown to stimulate insulin-dependent and –independent glucose uptake in cultured C2C12 muscle cells but not in 3T3-LI adipocytes [20], suggesting an insulin-mimetic and insulin-sensitizing effect in muscle. More recently, freeze-dried blueberry fruit supplementation (45g per day x 6 weeks) to obese insulin resistant individuals improved whole body insulin sensitivity (assessed by euglycaemic hyperinsulinaemic clamp technique) when compared to placebo [21]. Conversely high fat-fed mice provided freezedried whole blueberry powder (10% wt/wt in diet x 92 days) increased body weight and obesity, and did not alter glucose tolerance (assessed by IPGTT) [33]. There are a number of studies reporting on the effects of cinnamon on insulin sensitivity and glucose homeostasis (see review [30] and references therein). Cinnamon (20g/kg diet x 12 weeks) enhances whole body insulin sensitivity in high fat/sucrose fed rats without altering body weight or adiposity [31]. Similarly, cinnamon extract (300mg/kg/day in drinking water x 3 weeks) prevents insulin resistance in high fructose fed rats [32]. Numerous clinical studies have reported that cinnamon or cinnamon extract can improve glucose tolerance and/or whole body insulin sensitivity in healthy lean [34, 35] and insulin resistant [36] individuals. However the effect of these nutritional agents on insulin’s vascular and metabolic actions in muscle has not been previously explored.
Insulin-mediated microvascular recruitment in skeletal muscle plays an integral role in regulating muscle glucose uptake. We have shown that acute defects in insulin- mediated microvascular recruitment can contribute to the development of muscle insulin resistance in vivo. Infusion of α-methyl serotonin [26], endothelin-1 [27], L-NG-Nitroarginine methyl ester [7, 29, 37], TNFα [38], Intralipid® plus heparin [39] and glucosamine [40] in healthy animals attenuate insulin-mediated microvascular recruitment and insulin-mediated muscle glucose uptake. In addition, chronic models of insulin resistance including the high fat-fed rat (36% fat wt/wt) [3], and obese Zucker rat [41] display marked attenuation in insulin-mediated microvascular recruitment and muscle glucose uptake. More recently we reported that moderate increases in dietary fat cause microvascular and not myocyte-derived insulin resistance, which for the first time positioned defects in microvascular insulin action as an early event that contributes significantly to fat-induced muscle insulin resistance. In the present study, animals given BT for two weeks restored insulinmediated microvascular recruitment and this was associated with improved insulin-stimulated muscle glucose uptake, potentially due to improved delivery of insulin and glucose to myocytes.
This is the first report of an agent that reverses microvascular insulin resistance in muscle of the HFD rat model. Given that the HFD model we used in this study displays both myocyte and microvascular insulin resistance [14], the improvement in muscle glucose uptake following BT treatment could be due to improvements in both myocyte and microvascular insulin responsiveness. However, given that BT treatment reversed the impairment in microvascular insulin responsiveness and only ~40% of insulin-stimulated glucose uptake in muscle, it can be speculated that BT treatment has improved predominantly vascular and not myocyte insulin action. The current study also demonstrates that BT treatment markedly improved whole body, muscle and microvascular insulin sensitivity without changing adiposity or fasting insulin levels, two hallmarks of insulin resistance. This is suggestive of BT treatment having an insulin-sensitising action in this animal model, however whether these markers would be altered during longer term supplementation is unknown.
As detailed previously, BT is a blend of five plant based ingredients. The present study does not indicate which ingredient(s) or bioactive components of BT are responsible for this insulin sensitizing action, or whether in combination they have additive or synergistic actions. Although dried blueberries are the main constituent of this product, further studies using individual ingredients would be required to fully resolve these issues.
Conclusions
This is the first study to demonstrate that BT treatment markedly improves microvascular and metabolic insulin sensitivity in skeletal muscle of HFD rats. These effects were apparent independently of changes in adiposity, or clinical chemistries such as fasting plasma glucose and insulin. Our study suggests that regular consumption of BT may have important therapeutic implications for insulin resistance and T2D, however its efficacy in humans population remains to be determined.
Competing Interests
No conflicts of interest, financial or otherwise, are declared by the authors.
Authors’Contributions
D.P., K.M.R-T, M.A.K and were involved in design,conduct/data collection, analysis and writing of the manuscript. E.A.B., H.L.H.N, S.M.R. and S. R. were involved in design, analysis and writing of the manuscript. All authors read and approved the final manuscript.
Acknowledgements
This work was funded by the National Health & Medical Research Council of Australia (grant number 1009962 to M.A.K. and S.R.). Professor Stephen Rattigan is a Senior Research Fellow of the National Health & Medical Research Council of Australia (grant number 490034 to S.R). We thank Carl Sykes and Nadja Brearley of Dromana Blueberries for donating Blueberry BoostTM for this study.
References
- Coelho DF, Pereira-Lancha LO, Chaves DS, Diwan D, Ferraz R, Campos-Ferraz PL, Poortmans JR, Lancha Junior AH: Effect of high-fat diets on body composition, lipid metabolism and insulin sensitivity, and the role of exercise on these parameters. Braz J Med Biol Res 2011, 44:966-972.
- Kraegen EW, James DE, Storlien LH, Burleigh KM, Chisholm DJ: In vivo insulin resistance in individual peripheral tissues of the high fat fed rat: assessment by euglycaemic clamp plus deoxyglucose administration. Diabetologia 1986, 29:192-198.
- St-Pierre P, Genders AJ, Keske MA, Richards SM, Rattigan S: Loss of insulin-mediated microvascular perfusion in skeletal muscle is associated with the development of insulin resistance. Diabetes Obes Metab 2010, 12:798-805.
- Chadderdon SM, Belcik JT, Smith E, Pranger L, Kievit P, Grove KL, Lindner JR: Activity restriction, impaired capillary function, and the development of insulin resistance in lean primates. Am J Physiol Endocrinol Metab 2012, 303:E607-613.
- Defronzo RA, Gunnarsson R, Bjorkman O, Olsson M, Wahren J: Effects of insulin on peripheral and splanchnic glucose metabolism in noninsulin-dependent (type II) diabetes mellitus. JClinInvest 1985, 76:149-155.
- Rattigan S, Clark MG, Barrett EJ: Hemodynamic actions of insulin in rat skeletal muscle: evidence for capillary recruitment. Diabetes 1997, 46:1381-1388.
- Vincent MA, Clerk LH, Lindner JR, Klibanov AL, Clark MG, Rattigan S, Barrett EJ: Microvascular recruitment is an early insulin effect that regulates skeletal muscle glucose uptake in vivo. Diabetes 2004, 53:1418-1423.
- Vincent MA, Dawson D, Clark AD, Lindner JR, Rattigan S, Clark MG, Barrett EJ: Skeletal muscle microvascular recruitment by physiological hyperinsulinemia precedes increases in total blood flow. Diabetes 2002, 51:42-48.
- Vincent MA, Montagnani M, Quon MJ: Molecular and physiologic actions of insulin related to production of nitric oxide in vascular endothelium. CurrDiabRep 2003, 3:279-288.
- Zhang L, Vincent MA, Richards SM, Clerk LH, Rattigan S, Clark MG, Barrett EJ: Insulin sensitivity of muscle capillary recruitment in vivo. Diabetes 2004, 53:447-453.
- Baron AD: Hemodynamic actions of insulin. AmJPhysiol 1994, 267:E187-E202.
- Baron AD, Brechtel G: Insulin differentially regulates systemic and skeletal muscle vascular resistance. AmJPhysiol 1993, 265:E61-E67.
- Baron AD, Steinberg H, Brechtel G, Johnson A: Skeletal muscle blood flow independently modulates insulin-mediated glucose uptake. AmJPhysiol 1994, 266:E248-E253.
- Premilovac D, Bradley EA, Ng HL, Richards SM, Rattigan S, Keske MA: Muscle insulin resistance resulting from impaired microvascular insulin sensitivity in Sprague Dawley rats. Cardiovasc Res 2013, 98:28-36.
- Clerk LH, Vincent MA, Jahn LA, Liu Z, Lindner JR, Barrett EJ: Obesity blunts insulin-mediated microvascular recruitment in human forearm muscle. Diabetes 2006, 55:1436-1442.
- Keske MA, Clerk LH, Price WJ, Jahn LA, Barrett EJ: Obesity blunts microvascular recruitment in human forearm muscle after a mixed meal. Diabetes Care 2009, 32:1672-1677.
- Vincent MA, Clerk LH, Lindner JR, Price WJ, Jahn LA, Leong- Poi H, Barrett EJ: Mixed meal and light exercise each recruit muscle capillaries in healthy humans. AmJPhysiolEndocrinolMetab 2006,290:E1191-E1197.
- Eggleston EM, Jahn LA, Barrett EJ: Hyperinsulinemia rapidly increases human muscle microvascular perfusion but fails to increase muscle insulin clearance: evidence that a saturable process mediates muscle insulin uptake. Diabetes 2007, 56:2958-2963.
- DeFuria J, Bennett G, Strissel KJ, Perfield JW, 2nd, Milbury PE, Greenberg AS, Obin MS: Dietary blueberry attenuates whole-body insulin resistance in high fat-fed mice by reducing adipocyte death and its inflammatory sequelae. J Nutr 2009, 139:1510-1516.
- Martineau LC, Couture A, Spoor D, Benhaddou-Andaloussi A, Harris C, Meddah B, Leduc C, Burt A, Vuong T, Mai Le P, et al: Anti-diabetic properties of the Canadian lowbush blueberry Vaccinium angustifolium Ait. Phytomedicine 2006, 13:612-623.
- Stull AJ, Cash KC, Johnson WD, Champagne CM, Cefalu WT: Bioactives in blueberries improve insulin sensitivity in obese, insulinresistant men and women. J Nutr 2010, 140:1764-1768.
- Cignarella A, Nastasi M, Cavalli E, Puglisi L: Novel lipid-lowering properties of Vaccinium myrtillus L. leaves, a traditional antidiabetic treatment, in several models of rat dyslipidaemia: a comparison with ciprofibrate. Thromb Res 1996, 84:311-322.
- Allen FM: Blueberry Leaf Extract. Can Med Assoc J 1927, 17:1539- 1540.
- Kraegen EW, James DE, Jenkins AB, Chisholm DJ: Dose-response curves for in vivo insulin sensitivity in individual tissues in rats. Am- JPhysiol 1985, 248:E353-E362.
- Dawson D, Vincent MA, Barrett EJ, Kaul S, Clark A, Leong-Poi H, Lindner JR: Vascular recruitment in skeletal muscle during exercise and hyperinsulinemia assessed by contrast ultrasound. AmJPhysiolEndocrinolMetab 2002, 282:E714-E720.
- Rattigan S, Clark MG, Barrett EJ: Acute vasoconstriction-induced insulin resistance in rat muscle in vivo. Diabetes 1999, 48:564-569.
- Ross RM, Kolka CM, Rattigan S, Clark MG: Acute blockade by endothelin-1 of haemodynamic insulin action in rats. Diabetologia 2007, 50:443-451.
- Vincent MA, Barrett EJ: Insulin-induced capillary recruitment precedes changes in skeletal muscle glucose uptake. Diabetes2002,51:A31.
- Vincent MA, Barrett EJ, Lindner JR, Clark MG, Rattigan S: Inhibiting NOS blocks microvascular recruitment and blunts muscle glucose uptake in response to insulin. AmJPhysiolEndocrinolMetab 2003,285:E123-E129.
- Qin B, Panickar KS, Anderson RA: Cinnamon: potential role in the prevention of insulin resistance, metabolic syndrome, and type 2 diabetes. J Diabetes Sci Technol 2010, 4:685-693.
- Couturier K, Batandier C, Awada M, Hininger-Favier I, Canini F, Anderson RA, Leverve X, Roussel AM: Cinnamon improves insulin sensitivity and alters the body composition in an animal model of he metabolic syndrome. Arch Biochem Biophys 2010, 501:158-161.
- Qin B, Nagasaki M, Ren M, Bajotto G, Oshida Y, Sato Y: Cinnamon extract prevents the insulin resistance induced by a high-fructose diet. Horm Metab Res 2004, 36:119-125.
- Prior RL, Wu X, Gu L, Hager TJ, Hager A, Howard LR: Whole berries versus berry anthocyanins: interactions with dietary fat levels in the C57BL/6J mouse model of obesity. J Agric Food Chem 2008, 56:647-653.
- Solomon TP, Blannin AK: Effects of short-term cinnamon ingestion on in vivo glucose tolerance. Diabetes Obes Metab 2007, 9:895- 901.
- Solomon TP, Blannin AK: Changes in glucose tolerance and insulin sensitivity following 2 weeks of daily cinnamon ingestion in healthy humans. Eur J Appl Physiol 2009, 105:969-976.
- Wang JG, Anderson RA, Graham GM, 3rd, Chu MC, Sauer MV, Guarnaccia MM, Lobo RA: The effect of cinnamon extract on insulin resistance parameters in polycystic ovary syndrome: a pilot study. Fertil Steril 2007, 88:240-243.
- Bradley EA, Richards SM, Keske MA, Rattigan S: Local NOS inhibition impairs vascular and metabolic actions of insulin in rat musclein vivo. Am J Physiol Endocrinol Metab 2013.
- Youd JM, Rattigan S, Clark MG: Acute impairment of insulin-mediated capillary recruitment and glucose uptake in rat skeletal muscle in vivo by TNFa. Diabetes 2000, 49:1904-1909.
- Clerk LH, Rattigan S, Clark MG: Lipid infusion impairs physiologic insulin-mediated capillary recruitment and muscle glucose uptake in vivo. Diabetes 2002, 51:1138-1145.
- Wallis MG, Smith ME, Kolka CM, Zhang L, Richards SM, Rattigan S, Clark MG: Acute glucosamine-induced insulin resistance in muscle in vivo is associated with impaired capillary recruitment. Diabetologia 2005, 48:2131-2139.
- Wallis MG, Wheatley CM, Rattigan S, Barrett EJ, Clark AD, Clark MG: Insulin-mediated hemodynamic changes are impaired in muscle of zucker obese rats. Diabetes 2002, 51:3492-3498.





