Cellular Aging promotes CEMP1 Expression in STRO-1 positive PDL cells
Lamia AF1, Aminlari A2, Botero T2, Holland GR2, Kapila S3, Jin T4, Arzate H5, Kapila YL6*
1 Private Practice, Port Jefferson, New York, USA.
2 Department of Cariology, Restorative Sciences, and Endodontics, University of Michigan School of Dentistry, Ann Arbor, Michigan, USA.
3 Department of Orthodontics and Pediatric Dentistry, University of Michigan School of Dentistry, Ann Arbor, Michigan, USA.
4 Molecular Biology Core, University of Michigan School of Dentistry, Ann Arbor, Michigan, USA.
5 Faculty of Dentistry, National Autonomous University of Mexico,USA.
6 Department of Periodontics and Oral Medicine, University of Michigan School of Dentistry, Ann Arbor, Michigan, USA.
*Corresponding Author
Yvonne L. Kapila, DDS, PhD,
Professor, Department of Periodontics and Oral Medicine,
University of Michigan, School of Dentistry, 1011 North University Avenue,
Ann Arbor, MI 48109-1078, USA.
Tel: (734) 615-2295
E-mail: ykapila@umich.edu
Received: June 25, 2016; Accepted: July 13, 2016; Published: July 18, 2016
Citation: Kapila YL, Lamia AF, Aminlari A, Botero T, Holland GR, et al., (2016) Cellular Aging promotes CEMP1 Expression in STRO-1 positive PDL cells. Int J Dentistry Oral Sci.3(7), 276-280.DOI : dx.doi.org/10.19070/2377-8075-1600057
Copyright: Khalid K© 2016. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
We investigated the effects of cellular aging on the expression of cementum and mineralization-specific proteins in human periodontal ligament (PDL) cells, since this is not well studied. Previously, we showed that STRO-1- and cementum protein 1 (CEMP1) - positive PDL cells behave like cementoblasts, since they can be directed toward cementogenesis. However, the mineralization characteristics of these cells during aging have not been examined. Thus, we examined mineralization and expression of the mineralization-related proteins, CEMP1, collagen I (COL I) and osteopontin (OPN) in STRO-1 positive selected and unselected human PDL cells, control human cementoblastoma cells, and human mesenchymal stem cells (hMSC). Early (young) and late (aged) passage cells were compared. Aged STRO-1 positive cells exhibited the highest level of mineralization as assessed by Alizarin red staining and energy dispersive x-ray spectroscopy. Aged STRO-1 positive PDL cells also exhibited high levels of CEMP1, OPN, and COL I expression at two weeks compared to all other cells. At earlier time points, osteogenic induction significantly enhanced CEMP1 expression in aged STRO-1 positive and parental PDL cells. In contrast, STRO-1 negative cells exhibited high CEMP1 expression only in young cells. High CEMP1 expression was also present in young cells derived from cementoblastoma cells and hMSC. Expression of OPN and COL I varied for STRO-1 negative PDL, cementoblastoma, and hMSC cells. In summary, cellular aging significantly promotes mineralization and expression of CEMP1, OPN, and COL I in STRO-1 positive PDL cells. These findings may impact periodontal tissue function and regeneration in the context of aging tissues.
2.Introduction
3.Materials and Methods
3.1.Cell Culture
3.2.Magnetic Bead Cell Isolation
3.3.Immunofluorescence
3.4.Western Blot Analyses
3.5.Cellular Aging Protocol
3.6.Osteogenic/Cementogenic Differentiation
3.7.RNA extraction/Reverse Transcription
3.8.Quantitative Real-Time Polymerase Chain Reaction (qRTPCR) Assays
3.8.Alizarin Red S Staining
3.9.SEM and EDS
3.10.Statistical Analyses
4.Results
5.Discussion
6.Acknowledgements
7.References
Keywords
Cementum; Cementum Protein 1 (CEMP1); Periodontal Ligament Cells; Aging; Cementoblast; Senescence; Cementogenesis.
Introduction
The periodontium supports the tooth and consists of the periodontal ligament (PDL), tooth cementum, alveolar bone, and gingiva [1]. The PDL is comprised of a collagenous extracellular matrix and cells, including the cementum- producing cementoblasts [2]. Disease or injury to the periodontium can result from several factors [1, 3], however, effects of aging on the periodontium are not well known. Periodontal regeneration is difficult and requires reconstitution of several tissues, including root cementum, which is not predictable. Early steps in this process include attachment of new connective tissue fibers to root surfaces and cementogenesis. Cementogenesis is a critical process for homeostasis and regeneration of the periodontium [4, 5]. Although cementum was first described in 1835 and its recognition probably dates back to 1667 [6], it remains poorly understood at the cellular and molecular level.
Human cementoma-derived protein, cementum protein 1 (CEMP1) has been associated with cementogenesis [7, -12]. Transfection of CEMP1 into human gingival fibroblasts induced mineralization and expression of bone and cementum-matrix proteins [13, 14]. CEMP1 was expressed by cementoblasts and progenitor cells within the PDL. Since discovering residential stem cells in the PDL, recent studies have focused on their potential role in cementum formation. Indeed, PDL cells have the potential to differentiate into cementoblasts [15, -18]. Given these data, previous studies isolated postnatal stem cells from the PDL using the mesenchymal stem cell surface marker STRO-1 [15]. Paula-Silva and colleagues were the first to link STRO-1 and CEMP1 to cementogenesis [17]. Prior to this, molecules responsible for recruiting mesenchymal cells and inducing their differentiation into cementoblasts had not been identified. However, temporal expression and regulation of CEMP1 during mineralization and cellular aging are not known [7, - 10]. Cellular aging occurs as cells undergo multiple cell passages in culture [19, -23]. Thus, an in vitro cellular aging approach was employed in current studies to examine expression of mineralization related proteins, including CEMP1, in selected stromal cells.
Periodontal tissue breakdown and tooth loss increase with age. Studies have described changes of the PDL that occur with aging, however, these are largely limited to histological investigations [1, 2, 24, 25]. Therefore, understanding the biological changes of the PDL and its cells during the aging process could help shed light on the relationship between periodontal breakdown and aging. Previous studies proposed that CEMP1 is a mediator of periodontal regeneration and cementogenesis, since it stimulates PDL cell proliferation migration, and mineralization, and a cementum- like phenotype in gingival fibroblasts [13, 37, 17]. However, CEMP1 expression in PDL cells in relation to cellular aging has not been examined. Given an association between a STRO-1 positive phenotype and CEMP-1 expression in PDL cells with a regenerative potential [17], we selected cells expressing these markers to examine their contribution to cellular aging.
University of Michigan Institutional review board approval was obtained for this study. Premolars (n=24) extracted for orthodontic reasons were obtained from human subjects ages 12-16 and placed in culture media. PDL cells were harvested from teeth as previously described and cultured in basal α-minimal essential media (αMEM) containing 10% fetal bovine serum plus 1% penicillin/streptomycin [2], then selected by STRO-1 expression. Human cementoblastoma cells were obtained as previously described [7], and cultured in basal αMEM with 10% fetal bovine serum plus 1% penicillin/streptomycin. Human mesenchymal stem cells (hMSCs) and corresponding basal media were purchased (Lonza, MD, USA). All cells were prepared to passage 2 and 3 then used for experiments or aged as described below. Five cell types were used for these investigations: STRO-1 positive PDL cells, STRO-1 negative PDL cells, parental PDL cells, human cementoblastoma cells, and hMSC.
To isolate STRO-1 positive PDL cells, the CELLectin Biotin Binder Kit and a mouse anti- human STRO-1 antibody (1:20 dilution; R&D Systems Inc., Minneapolis, MN) and magnetic Dynabeads (Life Technologies, Grand Island, NY) were used with passage 3 (P3) PDL cells as previously described [26]. Bead bound cells were designated as STRO-1 positive cells and unbound cells were designated as STR0-1 negative cells. Parental PDL cells were the original starting cells from which STRO-1 positive and negative cells were derived.
Standard immunofluorescence was performed to confirm that magnetic bead isolated cells were STRO-1 positive. Since CEMP- 1 is a presumptive marker of cementoblasts [7 -14, 17], CEMP-1 expression was also examined in these cells. A mouse anti-human STRO-1 monoclonal IgM antibody (1:200 dilution; BioFx Labs, MD), and a CEMP1 antibody (1:300 dilution) [13, 14, 17] were used. Negative controls included using no antibody or a normal mouse IgG (1:150 dilution). A secondary FITC conjugated goat anti-mouse IgM antibody (1:300 dilution) and a Texas Red conjugated donkey anti-rat IgG antibody (1:300 dilution) were then applied. DAPI nuclear stain was used at a 1:5000 dilution. Cells were observed under light microscopy with fluorescence filters. Images were captured using Q-Capture Pro software and an Evolution MP digital camera attached to a Nikon TS100 microscope.
To confirm CEMP1 expression in isolated cells, standard western blot analyses were performed using an antibody to CEMP1 (described above) and an enhanced chemiluminescence detection system (West-Pico ECL, Pierce Biotechnology).
STRO-1 positive, STRO-1 negative, parental PDL, and positive control cementoblastoma cells were cultured in basal α-MEM and positive control hMSC cells were cultured in basal hMSC medium to 90% confluency (~2.5x106 cells). Cellular aging was performed by sub-cultivation according to Hayflick and Goseki [19, 20, 27]. Cells plated at P2 or P3 were either used immediately or passaged again for specific assays and considered ‘young’ cells (P3 - P4). For in vitro aging, cells were passaged and sub-cultivated using a 4:1 ratio. After P10 and P11, the growth rate of the cells decreased. The resulting cells were split into a 2:1 ratio to achieve passage 12 or more, and these late passage cells were considered ‘aged’ (P11-13).
Cells were incubated in osteogenic or control basal media for mRNA analysis, Alizarin Red S staining, and EDS analysis. Osteogenic induction media contained L-glutamine, ascorbate, dexamethasone, and β-glycerophosphate (Lonza, MD) [28]. RNA analysis was performed in a time course fashion (0h, 6h, 24h, 72h, 1wk, and 2wk) for both young and aged cells in the presence of osteogenic or control medium. Following the time course, samples underwent RNA extraction, reverse transcription and subsequent RT-qPCR.
RNA extraction was performed using the RNeasy Mini Kit (Qiagen, USA). RNA quality and concentration were evaluated on a Beckman DU-640 spectrophotometer using 260/280 absorbance. RNA samples underwent reverse transcription with the High Capacity cDNA Reverse Transcription Kit (Life Technologies, Carlsbad, CA).
qRT-PCR assays (30) were performed on quadruplicate cDNA samples using the TaqMan® Universal PCR Master Mix (Life Technologies, Carlsbad, CA) and the Vii7 Applied Biosystems PCR system (Applied Biosystems, Foster City, CA). TaqMan gene expression assay identification numbers and probe sequences for GAPDH, COL1A1, OPN, and CEMP1 are indicated in Table 1.


Table 1. TaqMan gene expression assay identification numbers and probe sequences for GAPDH, COL1A1, OPN, and CEMP1.
To assess mineralization, cells were incubated in osteoinduction or control media for 2 wk then stained and evaluated using the Alizarin red dye and cetylpyridinium chloride extraction method as reported previously [17]. Staining was quantified on a spectrophotometer at 570 nm. Images of samples were scanned using an Epson double-sided radiograph-scanning machine and an AppleMac computer.
To further assess mineralization, standard scanning electron microscopy (SEM) and energy dispersive x-ray spectroscopy (EDS) were performed as previously described [29]. Samples were washed in PBS, fixed in ice cold 70% ethyl alcohol and air dried prior to EDS and SEM analysis. Samples were covered with a thin (10nm) gold film then analyzed with a Leica-Cambridge 440 scanning electron microscope fitted with a PentaFET energy dispersive xray microanalysis microprobe. All analyses were performed at 20 kV for 300 sec.
Student’s t-tests were used for analysis to compare aged versus young cells. P < 0.05 was considered significant.
Results
To examine cellular aging on select PDL cells, STRO-1 positive PDL cells were first isolated and examined via immunofluorescence and western blotting (Figure 1). Since CEMP-1 is a presumptive marker of mineralization and cement genesis [17], its expression was also examined in these cells. STRO-1 positive selected PDL cells and positive control cementoblastoma cells consistently showed STRO-1 and CEMP1 expression in an overlapping pattern (Figure 1B). STRO-1 negative PDL cells failed to show comparable staining of these markers (Figure 1B). Western blot analyses confirmed that parental PDL cells, STRO-1 positive PDL cells, and control cementoblastoma cells all expressed CEMP1 (Figure 1C). However, STRO-1 negative PDL cells failed to express CEMP1. Combined immunofluorescence and western blot data indicate that bead isolation can be used to successfully identify STRO-1 positive, CEMP1 expressing cells from a parental pool of PDL cells.
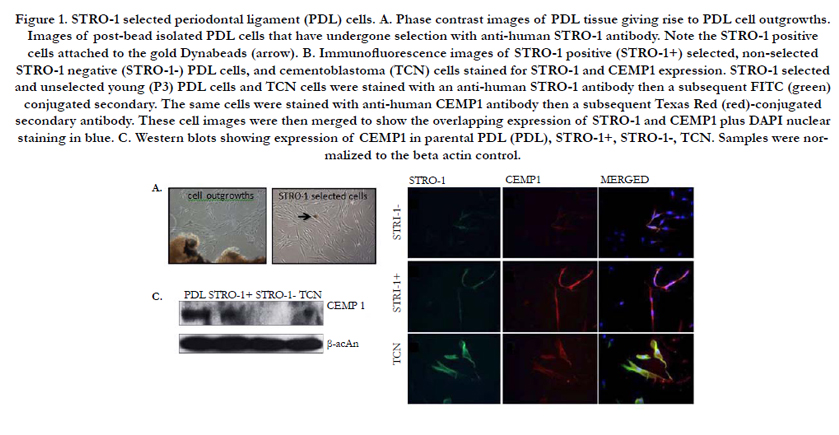
Figure 1. STRO-1 selected periodontal ligament (PDL) cells. A. Phase contrast images of PDL tissue giving rise to PDL cell outgrowths. Images of post-bead isolated PDL cells that have undergone selection with anti-human STRO-1 antibody. Note the STRO-1 positive cells attached to the gold Dynabeads (arrow). B. Immunofluorescence images of STRO-1 positive (STRO-1+) selected, non-selected STRO-1 negative (STRO-1-) PDL cells, and cementoblastoma (TCN) cells stained for STRO-1 and CEMP1 expression. STRO-1 selected and unselected young (P3) PDL cells and TCN cells were stained with an anti-human STRO-1 antibody then a subsequent FITC (green) conjugated secondary. The same cells were stained with anti-human CEMP1 antibody then a subsequent Texas Red (red)-conjugated secondary antibody. These cell images were then merged to show the overlapping expression of STRO-1 and CEMP1 plus DAPI nuclear staining in blue. C. Western blots showing expression of CEMP1 in parental PDL (PDL), STRO-1+, STRO-1-, TCN. Samples were normalized to the beta actin control.
Selected cells were next examined under osteogenic induction conditions. Images of cells revealed that osteogenic induction promoted nodule-like formation in parental PDL, STRO-1 positive, STRO-1 negative, and cementoblastoma cells to varying degrees in both young and aged cells (Supplemental Figure 1).
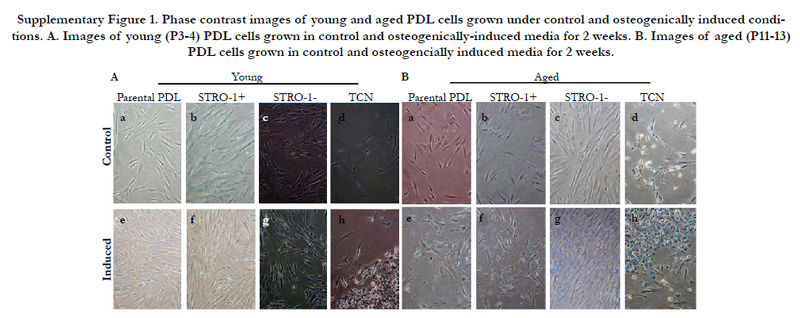

Supplementary Figure 1. Phase contrast images of young and aged PDL cells grown under control and osteogenically induced conditions. A. Images of young (P3-4) PDL cells grown in control and osteogenically-induced media for 2 weeks. B. Images of aged (P11-13) PDL cells grown in control and osteogencially induced media for 2 weeks.
To further assess mineralization potential, Alizarin red staining was evaluated in these cells (Figure 2). All young and aged STRO- 1 positive, parental PDL, and cementoblastoma cells exhibited significant Alizarin red staining under osteogenic treatment conditions. STRO-1 positive cells exhibited the highest level of Alizarin red staining compared to all cells under osteogenic conditions. STRO-1 negative PDL cells showed low levels of Alizarin red staining under osteogenic conditions (Figure 2). Interestingly, aged STRO-1 negative PDL cells exhibited higher Alizarin red staining than young STRO-1 negative cells under osteogenic conditions, suggesting that aging promotes mineralization even in cells not exhibiting the STRO-1 stemness marker (Figure 2).
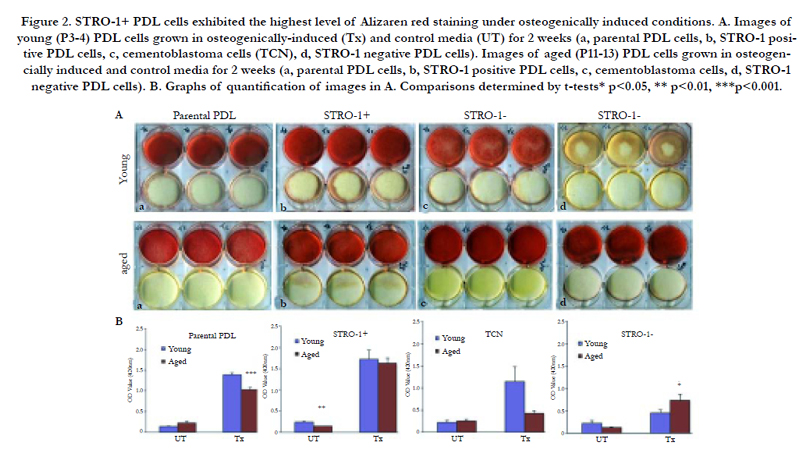
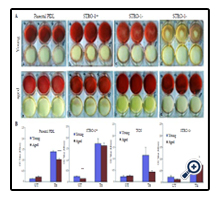
Figure 2. STRO-1+ PDL cells exhibited the highest level of Alizaren red staining under osteogenically induced conditions. A. Images of young (P3-4) PDL cells grown in osteogenically-induced (Tx) and control media (UT) for 2 weeks (a, parental PDL cells, b, STRO-1 positive PDL cells, c, cementoblastoma cells (TCN), d, STRO-1 negative PDL cells). Images of aged (P11-13) PDL cells grown in osteogencially induced and control media for 2 weeks (a, parental PDL cells, b, STRO-1 positive PDL cells, c, cementoblastoma cells, d, STRO-1 negative PDL cells). B. Graphs of quantification of images in A. Comparisons determined by t-tests* p<0.05, ** p<0.01, ***p<0.001.
EDS analysis revealed that the calcium to phosphate ratio (Ca/P) of mineral deposited by osteogenically induced cells was greatest for STRO-1 positive aged cells (Figure 3). STRO-1 positive aged cells exhibited Ca+2 and P energy peaks with an atomic percentage of 65.4 and 34.6, respectively, for a total 1.89 Ca/P. This value corresponds with that of biological hydroxyapatite [30]. Control hMSC exhibited Ca+2 and P energy peaks with an atomic percentage of 62.8 and 37.2, respectively, for a total 1.68 Ca/P. This value also corresponds well with that of biological hydroxyapatite. STRO-1 positive young cells revealed low peaks for Ca+2 and P (53.0 and 47, respectively) with a 1.13 Ca/P. Parental PDL cells showed even lower peaks for Ca+2 and P (52.2 and 47.7, respectively) with a 1.09 Ca/P. STRO-1 negative cells showed very low energy peaks for Ca+2 and P (23.9 and 76.0, respectively) with a 0.31 Ca/P. SEM images showed corresponding mineral deposition. The mineralization pattern for aged STRO-1 positive cells appeared distinct from that of other cells.
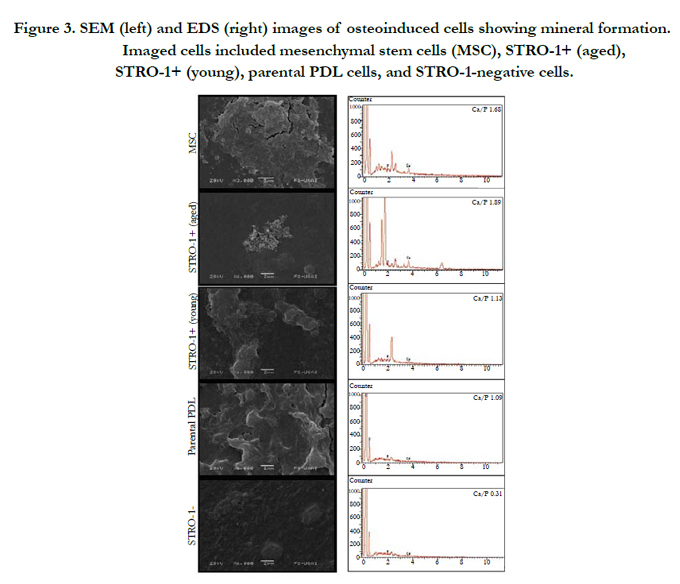
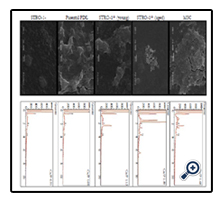
Figure 3. SEM (left) and EDS (right) images of osteoinduced cells showing mineral formation. Imaged cells included mesenchymal stem cells (MSC), STRO-1+ (aged), STRO-1+ (young), parental PDL cells, and STRO-1-negative cells.
The effect of in vitro cellular aging on the RNA expression of CEMP1 and other mineralization-related genes, osteopontin (OPN) and collagen I (COL I), was evaluated next using a time course approach and osteogenic or control media conditions. Fold increase and decrease RNA expression levels for CEMP1, OPN, and COL I are listed (Supplementary Tables I to IV). Evaluation of CEMP1 expression in aged parental PDL cells treated with and without osteogenic induction medium revealed a progressive increase in CEMP1 expression over time (Figure 4A). There was a 3-fold upregulation in CEMP1 expression for induced/treated and untreated aged cells at 24 h, a peak 7- fold upregulation for treated aged cells at 72 h, and a peak 12-fold upregulation at 1 wk for untreated aged cells (p<0.05). An increase in CEMP1 expression was also noted for aged STRO-1 positive PDL cells with a 2-fold and 5-fold upregulation at 1 wk for treated and untreated cells, respectively (p<0.05) (Figure 4B). CEMP1 expression for STRO-1 negative aged cells was low overall compared to all other cell types. A significant 3-fold downregulation for CEMP1 was noted for aged untreated STRO-1 negative cells at 24 h (p<0.05) and a 19-fold and 2.9-fold downregulation at 1 wk for both treated and untreated aged STRO-1 negative groups, respectively (p<0.05) (Figure 4C).
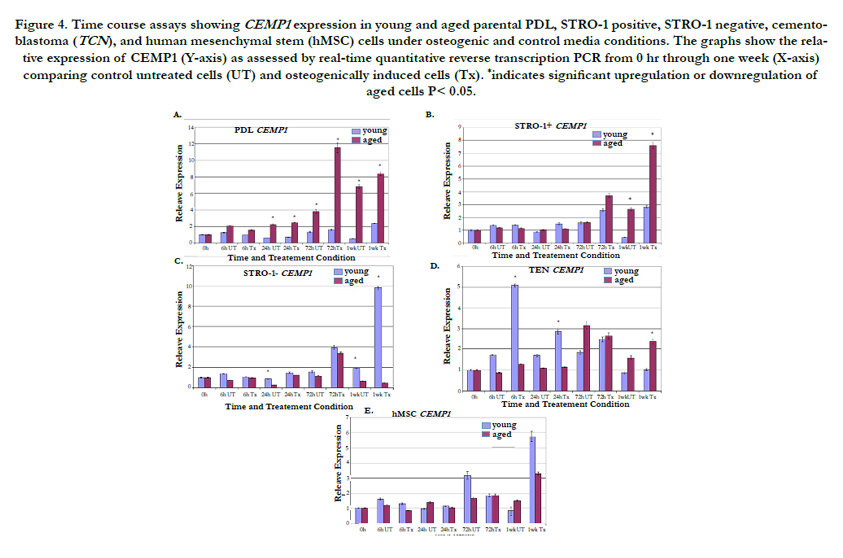
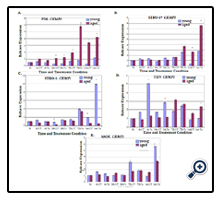
Figure 4. Time course assays showing CEMP1 expression in young and aged parental PDL, STRO-1 positive, STRO-1 negative, cementoblastoma (TCN), and human mesenchymal stem (hMSC) cells under osteogenic and control media conditions. The graphs show the relative expression of CEMP1 (Y-axis) as assessed by real-time quantitative reverse transcription PCR from 0 hr through one week (X-axis) comparing control untreated cells (UT) and osteogenically induced cells (Tx). *indicates significant upregulation or downregulation of aged cells P< 0.05.
Cementoblastoma cells exhibited a unique pattern of CEMP1 expression. Young cementoblastoma cells exhibited higher CEMP1 expression levels in earlier time points through 24 h. This pattern then switched, and aged cementoblastoma cells exhibited higher levels of CEMP1 expression at 1 wk (Figure 4D). HMSC exhibited elevated CEMP1 expression in young cells especially at later time points (Figure 4E).
Aged parental PDL cells showed a progressive increase in osteopontin expression with peak expression at 1 wk of treatment (Supplemental Figure 2A). STRO-1 positive aged cells also showed a progressive increase in osteopontin expression from 24 h to 1 wk with a peak 55-fold upregulation at 1 wk for induced/treated cells (p<0.05) as well as a significant upregulation at 1 wk for untreated control cells (p<0.015) (Supplemental Figure 2B). STRO-1 positive aged cells exhibited the highest level of osteopontin expression at 1 wk under treatment compared to all other cells. STRO-1 negative aged cells also showed increasing osteopontin expression over time under both treated and untreated conditions, except at 1 wk of treatment where young cells exhibited higher levels (Supplemental Figure 2C). Cementoblastoma cells showed mixed expression with peak levels noted at 72 h for both aged and young cells (Supplemental Figure 2D). Young hMSC exhibited increasing osteopontin expression that peaked at 1 wk (Supplemental Figure 2E).
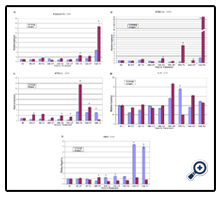
Supplementary Figure 2: Time course assays showing Osteopointin (OPN) expression in young and aged parental PDL, STRO-1 positive, STRO-1 negative, cementoblastoma (TCN), and human mesenchymal stem (hMSC) cells under osteogenic and control media conditions. The graphs show the relative expression of OPN (Y-axis) as assessed by real-time quantitative reverse transcription PCR from 0 hr through one week (X-axis) comparing control untreated cells (UT) and osteogenically induced cells (Tx). * indicates significant upregulation or downregulation of aged cells P< 0.05.
For collagen expression, aged parental PDL cells exhibited peak values at 1 wk. (Supplemental Figure 3A). STRO-1 positive aged PDL cells showed a progressive and significant upregulation of collagen I expression over time for all time points for both treated and untreated cells (Supplemental Figure 3B). Increased collagen I expression for STRO-1 positive aged cells began at 6 h and peaked at 1 wk, with an 18-fold increase in expression (p< 0.05). This collagen upregulation preceded the CEMP1 increased expression in these cells, suggesting that CEMP1 expression may be collagen dependent and important for late stages on mineralization. In contrast, young cells for the STRO-1 negative group showed the highest levels of collagen I expression (Supplemental Figure 3C). This collagen I expression peaked at 72 h then declined. Cementoblastoma cells showed peak collagen I expression at 24 h. Aged cementoblastoma cells showed higher collagen I expression under treatment conditions at 72 h and 1 wk, whereas young cells exhibited higher collagen I expression levels in the absence of treatment at 1 wk (Supplemental Figure 3D). Compared to young hMSC cells, aged hMSC cells consistently showed higher collagen I expression levels at all time points under treated or untreated conditions (Supplemental Figure 3E).
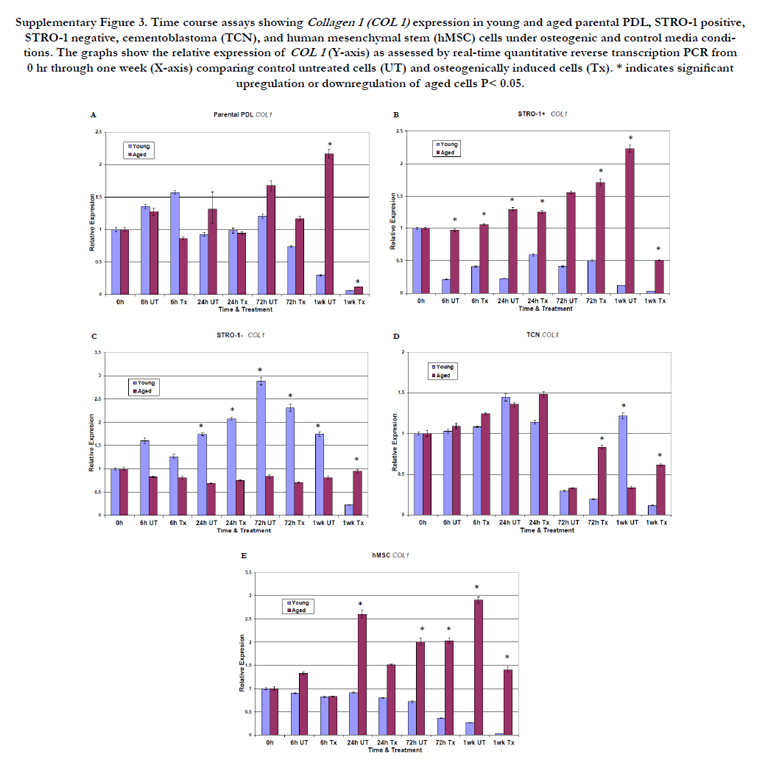

Supplementary Figure 3. Time course assays showing Collagen 1 (COL 1) expression in young and aged parental PDL, STRO-1 positive, STRO-1 negative, cementoblastoma (TCN), and human mesenchymal stem (hMSC) cells under osteogenic and control media conditions. The graphs show the relative expression of COL 1 (Y-axis) as assessed by real-time quantitative reverse transcription PCR from 0 hr through one week (X-axis) comparing control untreated cells (UT) and osteogenically induced cells (Tx). * indicates significant upregulation or downregulation of aged cells P< 0.05.
Examination of gene expression at 2 wk revealed a 208-fold upregulation of CEMP1 expression in aged STRO-1 positive PDL cells (P<0.05) (Figure 5A). There was a trend toward downregulation of CEMP1 expression in aged parental PDL, cementoblastoma, and STRO-1 negative cells, which was not statistically significant (P>0.05). In terms of collagen expression, aged STRO-1 positive cells again exhibited the highest levels of expression at the 2 wk mark (Figure 5B). Parental PDL cells and STRO-1 negative cells exhibited the next highest levels of collagen I expression at 2 wk. For osteopontin expression, STRO-1 positive cells again exhibited the highest levels along with parental PDL cells (Figure 5C).
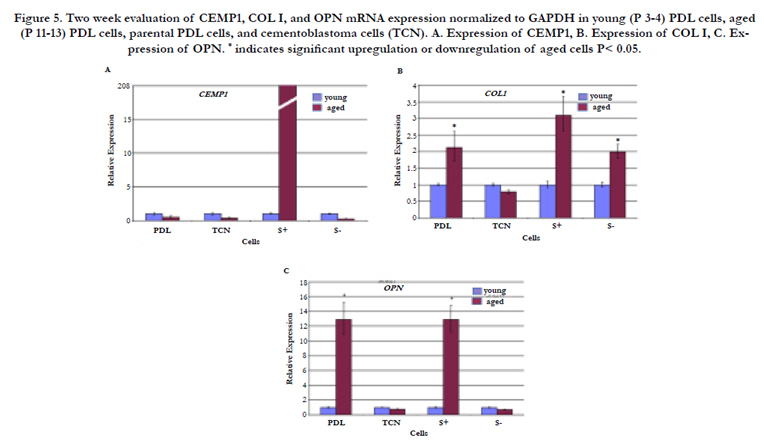

Figure 5. Two week evaluation of CEMP1, COL I, and OPN mRNA expression normalized to GAPDH in young (P 3-4) PDL cells, aged (P 11-13) PDL cells, parental PDL cells, and cementoblastoma cells (TCN). A. Expression of CEMP1, B. Expression of COL I, C. Expression of OPN. * indicates significant upregulation or downregulation of aged cells P< 0.05.
Discussion
Despite various methods and agents employed for periodontal regeneration [5, 31- 35], none have consistently promoted cementogenesis. Our work indicated that CEMP1 and STRO-1 may serve as markers of cementum forming cells or cementoblasts [17]. Cementoblasts are thought to produce cementum during regenerative processes [1, 4, 32, 33]. The PDL contains a subset of progenitor cells that are thought to give rise to cementoblasts [2, 24, 36]. These pluripotent cells have been isolated previously using the STRO-1 marker and magnetic beads as in the present study [37]. In current studies, we isolated STRO- 1 positive PDL cells to examine the effects of in vitro aging on CEMP1 temporal expression. This is the first report showing that CEMP1 expression increases with in vitro cellular aging. Expression of other mineralization associated genes, OPN and COL I, also increased significantly with cellular aging in these cells, indicating a general propensity toward mineralization with in vitro aging in pluripotent PDL cells. These data could have significant implications for periodontal tissue function and regeneration in the context of aging.
Osteogenic induction of aged STRO-1 positive cells promoted mineralization with features similar to that of biological hydroxyapatite [30]. However, the morphology of STRO-1 positive aged cells differed from that of other cells. These morphological differences could indicate alterations in underlying mineralization processes. In addition, the Ca/P ratio (1.89) of aged STRO-1 positive cells approximates that of cellular cementum (1.97) [9]. The hMSC Ca/P ratio (1.68) is similar to that of osteoblasts (1.628) [9]. These data suggest that aged STRO-1 positive PDL cells that express CEMP1 may be cementoblasts that can produce cementum-like mineral. The concept of CEMP1 regulating mineral deposition is not new [12, 17].
Cells aged in culture exhibit alterations in gene expression. Aged PDL cells exhibited decreased STRO-1, collagen XII and tenemodulin expression by P3, and further reduction in expression by P6 [22]. Similarly, in vitro aging of PDL cells and gingival fibroblasts up to P18 to P20 led to increased expression of COX- 2, IL-1 beta, IL-6, and tissue plasminogen activator [38, 39]. Our study showed that in vitro aging increased expression of CEMP1, OPN, and COL I in STRO-1 positive PDL cells and parental PDL cells. Increased CEMP1 expression seen in aging cells suggests a greater potential for cementum formation during cellular aging in a subset of progenitor PDL cells. Indeed the concept of increased cementum deposition with aging has been previously recognized [40].
In summary, we demonstrated that aging promotes mineralization in STRO-1 positive PDL cells. CEMP1 expression was significantly upregulated during the aging process, especially for STRO-1 positive cells. These findings enhance our knowledge of cementogenesis and cellular aging.
Acknowledgements
We would like to thank all members of the Kapila lab for their support and encouragement, especially Takayuki Hayami and Jinping Xu. The authors have no conflict of interest.
References
- Nanci A, Bosshardt DD (2006) Structure of periodontal tissues in health and disease. Periodontol 2000. 40: 11-28.
- Scanlon C, Marchesan J, Soehren S, Matsuo M, Kapila Y (2011) Capturing the regenerative potential of periodontal ligament fibroblasts. J Stem Cells Regen Med. 7(1):54-56.
- Engler WO, Ramfjord SP, Hiniker JJ (1966) Healing following simple gingivectomy. A tritiated thymidine radioautographic study. I. Epithelialization. J Periodontol. 37(4):298-308.
- Bosshardt DD, Schroeder HE (1996) Cementogenesis reviewed: a comparison between human premolars and rodent molars. Anat Rec 245(2):267- 292.
- Saito M, Iwase M, Maslan S, Nozaki N, Yamauchi M et al. (2001) Expression of cementum-derived attachment protein in bovine tooth germ during cementogenesis.Bone.29(3):242-248.
- Zander HA, Hürzeler B (1958) Continuous Cementum Apposition. J Dent Res. 37(6):1035-1044.
- Arzate H, Olson SW, Page RC, Narayanan AS (1992) Isolation of human tumor cells that produce cementum proteins in culture. Bone Miner 18(1): 15-30.
- Arzate H, Alvarez-Perez MA, Aguilar-Mendoza ME, Alvarez-Fregoso O (1998) Human cementum tumor cells have different features from human osteoblastic cells in vitro. J Periodontal Res. 33(5):249-258.
- Arzate H, Alvarez-Perez MA, Alvarez-Fregoso O, Wusterhaus-Chávez A, Reyes-Gasga J, Ximénez-Fyvie LA (2000) Electron microscopy, microanalysis,and X-ray diffraction characterization of the mineral-like tissue deposited by human cementum tumor-derived cells. J Dent Res. 79(1):28-34.
- Arzate H, Jimenez-Garcia LF, Alvarez-Pérez MA, Landa A, Bar-Kana I, Pitaru S (2002) Immunolocalization of a human cementoblastoma-conditioned medium-derived protein. J Dent Res. 81(8):541-546.
- Alvarez Perez MA, Pitaru S, Alvarez Fregoso O, Reyes Gasga J, Arzate H (2003) Anti- cementoblastoma-derived protein antibody partially inhibits mineralization on a cementoblastic cell line. J Struct Biol 143(1): 1-13.
- Alvarez-Perez MA, Narayanan S, Zeichner-David M, Rodríguez Carmona B, Arzate H (2006) Molecular cloning, expression and immunolocalization of a novel human cementum-derived protein (CP-23). Bone. 38(3):409-419.
- Carmona-Rodriguez B, Alvarez-Perez MA, Narayanan AS, Zeichner-David M, Reyes- Gasga J, et al., (2007) Human Cementum Protein 1 induces expression of bone and cementum proteins by human gingival fibroblasts. Biochem Biophys Res Comm. 358(3):763-769.
- Villarreal-Ramirez E, Moreno A, Mas-Oliva J, Chávez-Pacheco JL, Narayanan AS, Gil-Chavarría I, Zeichner-David M, et al., (2009) Characterization of recombinant human cementum protein 1 (hrCEMP1): primary role in biomineralization. Biochem Biophys Res Commun. 384(1):49-54.
- Seo BM, Miura M, Gronthos S, Bartold PM, Batouli S et al., (2004) Investigation of multipotent postnatal stem cells from human periodontal ligament. Lancet. 364(9429):149-155.
- Fuji S, Maeda H, Wada N, Tomokiyo A, Saito M, Akamine A (2008) Investigating a clonal human periodontal ligament progenitor/stem cell line in vitro and in vivo. J Cell Physiol. 215(3):743-749.
- Paula-Silva FW, Ghosh A, Arzate H, Kapila S, da Silva LA et al., (2010) Calcium hydroxide promotes cementogenesis and induces cementoblastic differentiation of mesenchymal periodontal ligament cells in a CEMP1- and ERK-dependent manner. Calcif Tissue Int. 87(2):144-157.
- Komaki M, Iwasaki K, Arzate H, Narayanan AS, Izumi Y, Morita I (2011) Cementum Protein 1 (CEMP1) induces a cementoblastic phenotype and reduces osteoblastic differentiation in periodontal ligament cells. J Cell Physiol. 227(2):649-657.
- Hayflick L, Moorhead PS (1961) The serial cultivation of human diploid cell strains. Exp Cell Res. 25:585-621.
- Hayflick L (1977) The cellular basis for biological aging: Handbook of the biology of aging. Van Nostrand Reinhold, New York. 159-179
- Ben-Porath I, Weinberg R (2005) The signals and pathways activating cellular senescence. Intl J Biochem Cell Biol. 37(5):961-976.
- Itaya T, Kagami H, Okada K, Yamawaki A, Narita Y et al., (2009) Characteristic changes of periodontal ligament-derived cells during passage. J Periodontal Res. 44(4):425-433.
- Rohme D (1981) Evidence for a relationship between longevity of mammalian species and life spans of normal fibroblasts in vitro and erythrocytes in vivo. Proc Natl Acad Sci USA. 78(8): 5009-5013.
- Lin NH, Menicanin D, Mrozik K, Gronthos S, Bartold PM (2008) Putative stem cells in regenerating human periodontium. J Periodontal Res. 43(5):514-523.
- Gomez Flores M, Hasegawa M, Yamato M, Takagi R, Okano T, Ishikawa I (2008) Cementum-periodontal ligament complex regeneration using the cell sheet technique. J Periodontal Res. 43(3): 364-371.
- Xu J, Wang W, Kapila Y, Lotz J, Kapila S (2009) Multiple differentiation capacity of STRO-1+/CD146+ PDL mesenchymal progenitor cells. Stem Cells Dev.18(3):487-496.
- Goseki T, Shimizu N, Iwasawa T, Takiguchi H, Abiko Y (1996) Effects of in vitro cellular aging on alkaline phosphatase, cathepsin activities and collagen secretion of human periodontal ligament derived cells. Mech Ageing Dev. 91(3):171-183.
- Hayami T, Zhang Q, Kapila Y, Kapila S (2007) Dexamethasone's enhancement of osteoblastic markers in human periodontal ligament cells is associated with inhibition of collagenase expression. Bone. 40(1):93-104.
- Saito E, Saito A, Kawanami M (2003) Favorable healing following space creation in rhBMP-2-induced periodontal regeneration of horizontal circumferential defects in dogs with experimental periodontitis. J Periodontol.74(12):1808-1815.
- Van den Bos T, Handoko G, Niehof A, Ryan LM, Coburn SP et al., (2005) Cementum and dentin in hypophosphatasia. J Dent Res. 84(11):1021– 1025.
- Heijl L, Heden G, Svärdström G, Ostgren A (1997) Enamel matrix derivative (EMDOGAIN) in the treatment of intrabony periodontal defects. J Clin Periodontol. 24(9):705-714.
- Krebsbach PH, Robey PG (2002) Dental and skeletal stem cells: Potential cellular therapeutics for craniofacial regeneration. J Dent Educ. 66(6):766- 773.
- Saygin NE, Giannobile WV, Sommerman MJ (2000) Molecular and cell biology of cementum. Periodontol. 24:73-98.
- Yoshikawa G, Murashima Y, Wadachi R, Sawada N, Suda H (2002) Guided bone regeneration (GBR) using membranes and calcium sulphate after apicectomy: a comparative histomorphometrical study. Int Endod J.35(3):255-263.v
- Bosshardt DD, Sculean A, Donos N, Lang NP (2006) Pattern of mineralization after regenerative periodontal therapy with enamel matrix proteins.Eur J Oral Sci. 114 (1):225-231.
- McCulloch CA (1985) Progenitor cell populations in the periodontal ligament of mice. Anat Rec. 211(3):258-262.
- Xu J, Wang W, Kapila Y, Lotz J, Kapila S (2009) Multiple differentiation capacity of STRO-1+/CD146+ PDL mesenchymal progenitor cells. Stem Cells Dev.18(3):487-496.
- Abiko Y, Shimizu N, Yamaguchi M, Suzuki H, Takiguchi H (1998) Effect of aging on functional changes of periodontal tissue cells. Ann Periodontol 3(1): 350-369.
- Shimizu N, Goseki T, Yamaguchi M, Iwasawa T, Takiguchi H et al., (1997) In vitro cellular aging stimulates interleukin-1 beta production in stretched human periodontal-ligament-derived cells. J Dent Res. 76(7): 1367-1375.
- Stein TJ, Corcoran JF (1990) Anatomy of the root apex and its histologic changes with age. Oral Surg Oral Med Oral Patho 69(2):238-242.





