The Expression of Platelet-Activating Factor is Induced by Low Extracellular Mg2+ in Aortic, Cerebral and Neonatal Coronary Vascular Smooth Muscle; Cross Talk with Ceramide Production, NF–kB and Proto-Oncogenes: Possible Links to Atherogenesis and Sudden Cardiac Death in Children and Infants,and Aging; Hypothesis, Review and Viewpoint
Altura BM1,2,3,4,5*, Li W1, Zhang A1, Zheng T1, Shah NC1, Gatha J. Shah1, Pérez-Abela JL6, Altura BT1,3,4,5
1 Department of Physiology & Pharmacology, SUNY Downstate Medical Center, Brooklyn, NY, USA.
2 Department of Medicine, SUNY Downstate Medical Center, Brooklyn, NY, USA.
3 Center for Cardiovascular and Muscle Research, SUNY Downstate Medical Center, Brooklyn, NY, USA.
4 The School of Graduate Studies in Molecular and Cellular Science, State University of New York Downstate Medical Center, Brooklyn, NY, USA.
5 Bio-Defense Systems, Inc, Rockville Centre, NY, USA.
6 Instituto Bien de Salud, Lima, Peru.
*Corresponding Author
Burton M. Altura,
Professor, Department of Physiology and Pharmacology,
SUNY Downstate Medical Center, Brooklyn, NY 11203, USA.
Tel: 718-270-2194
E-mail: baltura@downstate.edu
Received: February 29, 2016;Accepted: March 22, 2016;Published: March 31, 2016
Citation: Altura BM et al., (2016) The Expression of Platelet-Activating Factor is Induced by Low Extracellular Mg2+ in Aortic, Cerebral and Neonatal Coronary Vascular Smooth Muscle; Cross Talk with Ceramide Production, NF –kB and Proto-Oncogenes: Possible Links to Atherogenesis and Sudden Cardiac Death in Children and Infants, and Aging; Hypothesis, Review and Viewpoint. Int J Cardiol Res. 3(1), 47-67.DOI :dx.doi.org/10.19070/2470-4563-1600011
Copyright: Altura BM© 2016. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, pro/vided the original author and source are credited.
Abstract
An attempt is made, herein, to reconcile, and integrate, various phenomena associated with magnesium deficiency (MgD) in cardiovascular health, disease, and aging as well as reasons for the high incidence of sudden cardiac death in infants and young adults. With new experiments, we demonstrate, for the first time, that very low concentrations of platelet-activating factor (PAF), when added to primary cultured cerebral, neonatal coronary, and aortic vascular smooth muscle (VSM) cells (from three different mammals) promote rapid rises in free intracellular Ca2+ ions and a significant, concomitant reduction in free intracellular Mg2+ ions; these actions of PAF being curtailed with a specific membrane-receptor inhibitor of PAF. Our new experiments also demonstrate that addition of PAF to the VSM cells result in activation of NF-kB, activation of the proto-oncogenes c-fos and c-jun, a generation/release of ceramide, and synthesis of DNA; most of these actions being inhibited by a specific membrane-receptor antagonist of PAF. In addition, we show, for the first time, formation of 4-hydroxy-2-nonenal (4-HNE) in VSM cells incubated in Mg-deficient (MgD) environments or after addition of PAF. This is important because 4-HNE is a well-known inducer of hydrogen peroxide, known to be formed in MgD VSM cells and that 4-HNE is known to induce DNA damage and fragmentation, events that we have shown previously in VSM and cardiac muscle cells exposed to low Mg environments. Lastly, our experiments demonstrate that incubation of cerebral, neonatal coronary and aortic VSM cells in low Mg2+ induces: 1. rapid formation of PAF which can be attenuated greatly with a specific membrane-receptor antagonist of PAF; and 2. rises in cellular levels of ceramide, NF-kB activation and formation of the proto-oncogenes, and synthesis of DNA, all of which could be inhibited with a specific membrane-receptor antagonist of PAF. These new findings suggest major roles for PAF (and probably PAF-like lipids) and ceramide formation in the cardiovascular manifestations of MgD, inflammation, atherogenesis, aging, and sudden cardiac death in children. The significance of our new findings are reviewed in light of other works on MgD, inflammation, atherogenesis, sudden cardiac death, and aging.
2.Introduction
3.Materials and methods
3.1 Animals, tissue preparation and incubation
3.2 Primary culture of aortic and cerebral VSM cells
3.3 [Ca2+]I measurements
3.4 [Mg2+]I measurements
3.5 Expression of NF-kB
3.6 Induction and measurement of c-fos and c-jun
3.7 Measurement of cellular levels of ceramide
3.8 DNA synthesis
3.9 Measurement of PAF
3.10 Measurement of 8-hydroxydeoxyguanosine (8-OH-dg)
3.11 Measurement of protein-bound 4-HNE( 4-hydroxy-2-nonenal)
3.12 Chemicals and drugs
3.13 Statistics
4.Results
4.1 Influence of [Mg2+]0 on [Ca2+]i :effects of WEB2086
4.2 Influence of PAF on [Ca2+]I and [Mg2+]i
4.3 Low [Mg2+]0 and PAF result in early expression of proto-oncogenes : effect of WEB2086
4.4 Exposure of primary cultured VSM cells to low [Mg2+]0 or PAF results in NF-kB p65 expression: influence of PAF inhibitor
4.5 Exposure of primary cultured VSM cells to low [Mg2+]0 or PAF results in generation/release of ceramide: influence of WEB2086 , desipramine and scyphostatin
4.6 Influence of PAF and low Mg2+ on 4-HNE levels: Effects of WEB2086
5.Discussion
5.1 Molecular pathways for synthesis of ceramides and low Mg
5.2 Low Mg ,intracellular Ca, PAF,PAF-like lipids and ceramide
5.3 Significance of low Mg-induced DNA synthesis and DNA oxidation: roles of PAF, oxidized phospholipids, and ceramide
5.4 Mechanisms by which PAF and PAF-Rs function in VSM cells: roles of NF-kB, proto-oncogenes, Mg2+, phenotypic changes and vascular remodeling
5.5 Magnesium deficiency, PAF and unexplained sudden cardiac deaths in infants, children and young adults
5.6 Low Mg, PAF , telomerase activity , and aging
5.7 The microcirculation, white blood cells ,inflammation, atherogenesis , MgD , PAF and ceramide
5.8 MgD, PAF, N-SMases, acid-SMAses, ceramide, and Ca2+
5.9 Magnesium deficiency and genotoxicity
5.10 Low Mg intake and epigenetic processes
5.11 Bioavailable Mg intake and prevention of cardiovascular diseases
6.Acknowledgements
7.References
KeyWords
platelet-activating factor; Neutral -sphingomyelinase; Acid sphingomyelinase; PAF-like lipids; C-fos; C-jun; 8-hydroxydeoxyguanosine; 4-hydroxy-2-nonenal (4-HNE).
Introduction
A number of physiologic, nutritional, and biochemical homeostatic control mechanisms are responsible for normal functions of the peripheral, coronary and cerebral vasculatures. These homeostatic factors maintain both the patency and the fluid of the blood and its elements contained in them. Pathologic changes in the peripheral, coronary and cerebral vasculatures can result in alterations in vascular wall geometry, arterial and endothelial surfaces and in disturbances in oxygenation and nutritional status of blood vessels and the tissues they perfuse. Although many theories and hypotheses have been generated to account for hypertrophy and stiffening of resistance vessels in the etiology of hypertension, there is no agreement as to the precise mechanisms [for reviews, see 1, 2].
Approximately 35 years ago, our laboratory suggested that a progressive, dietary and/or a metabolic-induced loss of magnesium [Mg] from the body beginning early in life, particularly in and around the coronary arteries , could lead to coronary arterial spasm, ischemic heart disease , and sudden cardiac death [3]. Ever since this work was published, a number of clinical studies have been done and published which support this hypothesis, at least in adults [4-11]. However, little or no studies have been published to either confirm or deny this hypothesis in either infants or young adults. Autopsies of children , who have died as a consequence of accidents, have demonstrated early signs of atherogenesis (i.e., fatty streaks on the walls of the aorta and carotid arteries in young children as early as six years of age)[12]. It should be noted that atherosclerosis is the number- one cause of premature death in developing countries, including the United States and plays a major role in etiology of hypertension and strokes.
Disturbances in diet are known to produce inflammatory lesions, promote lipid deposition and accelerated growth and transformation of the smooth muscle cells in the vascular walls [for reviews, see 13-18]. Reduction in dietary magnesium intake has been demonstrated, experimentally, to result in hypertension [4, 5,17-20], atherogenesis [16,21-27], and stroke[15,27-34]. Hypermagnesemic diets have been shown to ameliorate hypertension, atherogenesis, stroke and certain inflammatory responses [14-18, 20-22,24,25,28,29,31,34-41]. In the Western World, dietary intake of magnesium is subnormal, with shortfalls of between 65 and 275 mg of Mg/d, depending on geographic region [14, 42,43]. Newly compiled USA HHANES data indicate that approximately 65 % of the American population is Mg deficient [44]. Several epidemiologic studies in North America and Europe have shown that children and adults consuming Western-type diets are low in Mg content as are pregnant women (i.e., < 30-50 % of the RDA for these populations) [15,43-47].
Using sensitive, specific Mg2+- ion selective electrodes, it has been shown that patients with hypertension, ischemic heart disease, and stroke exhibit significant depletion of serum and cellular ionized Mg2+ , the physiologically active Mg [15,25,29-31,35,57,71] . Such low levels of ionized Mg2+ , when mimicked in vitro, result in spasms of peripheral, coronary, placental, neonatal, and cerebral vascular smooth muscle cells, as well as increasing vascular reactivity to numerous neurogenic and humoral molecules [3-5,15,19, 25-31, 45-58, 72]. Dietary deficiency of Mg in rats has been shown to not only cause hypertension, but to also cause vascular remodeling (i.e., arteriolar wall hypertrophy of unknown origin), rarefaction of capillaries (known in itself to result in hypertension), as well as release of cytokines and chemokines( involved in inflammatory and atherogenic responses)23,74-82].
The nuclear factor-kappa B (NF-kB) and the proto-oncogenes (e.g., c-fos, c-jun) are two major regulators of growth, differentiation, cell migration, and cell death (e.g., apoptosis)[for reviews, see 83-86]. NF-kB is a transcription factor and a pleiotropic regulator of numerous genes involved in inflammatory responses, hypertension and atherogenesis [for reviews, see 83-85, 87]. Both NF-kB and the proto-oncogenes are thought to be important in numerous vascular disease processes such as inflammation, atherogenesis, hypertension, and stroke-like events [for reviews, see 83-87]. It is, however, not clear as to what initiates expression of these molecular and cellular events, particularly with respect to how Mg deficiency impacts on these vascular events and how low Mg causes hypertension, intense vasospasm, inflammation, stroke-like events, cardiac failure, and sudden cardiac death. Experiments performed in -vitro and in-vivo have demonstrated that short-term magnesium deficiency (MgD) results in formation/activation of NF-kB and the proto-oncogenes in cardiovascular tissues and VSM cells [15, 25,26,29,81,82,88].
Recently, we have provided experimental evidence for a role for sphingolipids, and particularly generation/release of ceramide, in initiating the sequelae of vascular events brought about via Mg deficiency [15, 25, 81, 89-99]. We have found, in cardiovascular tissues and diverse cell types, that both Mg deficient diets and in-vitro low extracellular Mg ions upregulate all three major, enzymatic biochemical pathways causing increased levels of cellular ceramide, i.e., upregulation of serine palmitoyl transferase 1 and 2, sphingomyelinases(N-, acid-, and alkaline- SMases),and ceramide synthase[15, 81, 82, 90, 93-95, 97, 98]. More recently, we have identified the existence of a fourth pathway for synthesis of ceramide in diverse cardiac and VSM cells, namely , "the salvage pathway" discovered by Merrill and co-workers in other cell types [98] In these studies, we also demonstrated that activation of protein kinase C isozymes play major roles in generation of ceramides [98]. We found these sphingolipid pathways play a major role in Mg-deficient upregulation of proto-oncogene (c-fos, c-jun) expression, NF-kB expression as well as DNA synthesis, protein kinase C families of enzymes, cytokines, and telomerase activity in diverse cardiac and vascular smooth muscle (VSM) cells [15,81,82,89-91,93-95,97-99].
Telomeres are critical in regulating genome integrity and hence chromosomal integrity [100, 101]. Telomeric DNA characteristics are highly conserved molecules among all eukaryotes [100, 101]. The telomeres are found at the ends of the chromosomes, consisting of short nucleotide repeats and specialized proteins ,which are regulated by telomerases [100,101]. Chromosomes devoid of telomeres or low telomerase activities undergo apoptosis [102-104]. Thus, cells with normal telomerase activities are thought to survive periods of oxidative stress (e.g., production of reactive oxygen and nitrogen species) and curtail apoptosis and the aging process [100-104]. Since MgD states have recently been shown, both in vivo and in vitro , to promote apoptosis and oxidative stress(i.e., DNA damage and production of reactive oxygen and nitrogen species) as well as downregulation of telomerase activities , in diverse cardiovascular tissues and VSM cells, we hypothesized that prolonged MgD would lead to premature aging [99]. During these Mg deficient experiments, we noted a fairly rapid synthesis of platelet-activating factor (PAF; 1-O-alkyl-2-acetyl-sn-glycero-3-phosphocholine)[unpublished findings]. In 1997, using primary culture of rat aortic and canine cerebral VSM cells and proton nuclear magnetic resonance spectroscopy, we saw peaks in the spectra that suggested that Mg - deficient environments led to rapid PAF and PAF-like molecules [89].
PAF is manufactured from a specific subclass of phosphatidylcholines which contain an ether, instead of an ester, bond at the sn-1 position of the glycerol backbone. This phospholipid precursor is a minor part of low density lipoprotein (LDL), which we and others have found to be elevated in blood and tissues of magnesium deficient animals [91,105-108]. LDL is a major component in a few blood cells, such as leukocytes, and along with ox-LDL, is a major pro-atherogenic substance [109,110]. A polyunsaturated fatty acid like arachidonate is found in the sn-2 position in the subclass of phosphatidylcholine molecules, and hydrolysis of this precursor phospholipid forms arachidonate for eicosanoid syntheses [110]. Acetylation of the other reaction product, viz., alkyl lysophosphatidylcholine, with acetyl-CoA finalizes the synthesis of PAF, both enzymes being tightly regulated [110]. It is important to note that because the precursors of PAF contain polyunsaturated fatty acids, they are very susceptible to oxidative attack, often resulting in a number of structural products that resemble PAF ( i.e., PAF-like lipids) [110]. Interestingly, approximately 20 years ago we reported , using proton-NMR spectroscopy and primary cell culture of diverse VSM cells, that lowering [Mg2+]0 in the culture media resulted in changes in phosphatidylcholine levels and production of several PAF-like molecules [89], most of which can activate the PAF receptors by a G1-coupled mechanism [110-114].
Since PAF is now thought to be an important, maybe critical, molecule in etiology of atherosclerosis and several inflammatory conditions and diseases( i.e., allergies, asthma, sepsis, acute pancreatitis, diverse bowel disorders, pulmonary diseases, diabetes, trauma, and shock among others)[110-115], we designed experiments with primary cell cultures of adult rat aortic, piglet coronary arterial, and adult canine cerebral VSM cells to determine what role(s), if any, PAF might play in the generation/release of the previously observed increased generation/release of cellular ceramide, proto-oncogene activation, activation of NF-kB, and DNA synthesis and oxidation. Since reactive oxygen species ( which are formed in MgD environments) can directly cause covalent modifications in DNA [115-119], we also investigated whether low [Mg2+ ]0 result in activation of stress-signaling pathways by examination of production of 4-hydroxy-2-nonenal(HNE). HNE, a major aldehyde product of lipid peroxidation in membranes, is known to exert numerous cytotoxic, genotoxic, biological, and signaling actions [116-119]. As little as 1.0 uM of HNE can produce chromosomal abnormalities and result in DNA fragmentation. We, thus, hypothesized that both PAF and low [Mg2+]0 would result in production of HNE in cultured cerebral, aortic and neonatal coronary arterial VSM cells.
Since previous data indicates that low Mg environments can, paradoxically, both induce synthesis of DNA and promote DNA damage in primary cultured aortic and cerebral VSM cells [26,91,99], an attempt is made to reconcile, and integrate, these phenomena with PAF activation in atherogenesis, inflammation and aging in order to possibly explain many of the cardiovascular effects of MgD.
We demonstrate, herein, for the first time, that very low concentrations of PAF when added to primary cultured cerebral, neonatal coronary and aortic VSM cells (from three different mammals) promote rapid rises in free intracellular Ca2+ ions ([Ca2+]i) and a significant, concomitant reduction in free intracellular Mg2+ ions ([Mg2+ ]i ); these actions of PAF being curtailed with a specific membrane -receptor inhibitor of PAF. Our new experiments also demonstrate that addition of PAF to the VSM cells result in activation of NF-kB , activation of the proto-oncogenes c-fos and c-jun, a generation/release of ceramide, and synthesis of DNA; most of these actions being inhibited by a specific membrane-receptor antagonist of PAF. In addition, we demonstrate, for the first time, formation of HNE in VSM cells incubated in MgD environments. This finding is important because HNE is a well-known inducer of hydrogen peroxide, formed in MgD VSM cells [15,88,unpublished findings], and that HNE is known to induce DNA damage and fragmentation, events that we have shown are also found in VSM and cardiac muscle cells exposed to low Mg environments. Lastly, our experiments show that incubation of the cerebral, neonatal coronary and aortic VSM cells in low [Mg2+]0 induce: 1. rapid formation of PAF which can be attenuated greatly with an inhibitor of PAF ; and 2. rises in cellular levels of ceramide, NF-kB activation and formation of the proto-oncogenes, and synthesis of DNA, all of which could be inhibited greatly with a specific membrane-receptor antagonist of PAF. These new findings suggest major roles for PAF (and probably PAF-like lipids) and ceramide formation in the cardiovascular manifestations of MgD , inflammation , atherogenesis, aging and sudden cardiac death in children. The significance of our new findings are reviewed in light of other works on MgD , inflammation, atherogenesis, sudden cardiac death and aging.
Materials and methods
Adult male rats (16 to 20 weeks old, 275-325 g) were killed by decapitation and exsanguinated as previously [26, 30,51]. All experiments were approved by The Animal Use and Care Committee of the State University of New York Downstate Medical Center. Thoracic aortae were excised, immediately placed in normal Krebs-Ringer bicarbonate (NKRB) solution at room temperature, cleaned of blood, loose connective tissue, and fat, and the aortae cut into rings about 2 to 3 mm long, as described previously [26,30,51]. Endothelial cells were rubbed off with wet filter paper to get only pure, viable VSM cells [51, 81,82, 85]. Concentrations of ionized Mg2+ and Ca2+ in culture media were determined precisely using ion-selective electrodes (NOVA Biomedical Corp., Waltham ,MA)[57].
Mongrel dogs (15+/- 3 kg) were anesthetized with intravenous sodium pentobarbital (40 mg/kg) and sacrificed by bleeding from the common femoral arteries [30]. After craniotomy, the brains were rapidly removed, placed in normal NKRB solution at room temperature, and basilar and middle cerebral arteries excised, cleaned of arachnoid membranes and blood elements, and the vessels were denuded of endothelium as described previously[30,48]. The vessels were cut into segments about 3 to 4 mm in length [48].
Four-five piglets (7-10 days old) were anesthetized with saffan (12 mg/kg i.m.; Glaxo or Pittman Moore Cos.). The hearts were removed and coronary arteries were extracted and denuded of endothelium, as previously [120], prior to enzymatic digestion. By use of monoclonal antibodies for alpha-actin and trypan blue exclusion, we found that 96-98 % of the VSM cells were pure smooth muscle cells [120].The pure VSM cells were cultured as described previously, in Dulbecco's modified Eagle's media, in a humidified atmosphere composed of 95% air-5 % CO2 [120] . The cells were exposed to media containing either 0.15, 0.30, 0.48, 0.6, or 1.2 mM [Mg2+]0 for periods of from 15 min to 18-24 hr prior to manipulations.
Rat aortic and canine cerebral VSM cells were isolated according to established methods in our labs and cultured in Dulbecco’s modified Eagle’s medium containing 1.2 mmol/l [Mg2+]0 , fetal calf serum (FCS) and antibiotics at 37°C in a humidified atmosphere composed of 95% air-5% CO2 [26,30,51]. After confluence was reached, the VSM cells were placed in media containing either 0.15, 0.30, 0.6, 1.2, or 4.8 mmol/l [Mg2+]0 for varying periods of time ( 15 to 120 min or 18 to 20 hr) .
Intracellular free cytosolic Ca2+ ([Ca2+]i) was measured with digital image analysis and fura-2/AM as detailed previously [26,30,51,55]. Primary smooth muscle cells obtained from rats , dogs and piglets (as in above) were seeded on glass cover slips (12 mm in diam; about 1x 106 cells per cover slip) and used 2 to 3days after seeding as described previously [26,30,51,55]. After exposing the cells to either vehicle(NKRB) with and without PAF or NKRB with varying [Mg2+]0 with and without PAF, monolayers of VSM cells , grown on the coverslips, were loaded with 2.0 umol/l fura-2 AM and 0.12 % pluronic acid F-127 (60 min, 37°C)[26,30,51,55]. In some experiments, we added the PAF inhibitor, WEB2086 (5uM, Sigma Biochemicals, St. Louis, MO), scyphostatin (75 uM; N-SMAse inhibitor),or desipramine (20 uM; acid-SMAse inhibitor) to the incubation media. The experimental procedures for [Ca2+]I measurements were carried out as described previously with the use of fura-2 AM [26,30,51,55]. The [Ca2+]I was calculated according to the following equation:
A Kd of 224 nmol/l was used for the fura-2/Ca2+: fura-2 complex excited at 380 nm
Intracellular free cytosolic Mg2+ ([Mg2+ ]i) was measured with digital image analysis and mag fura-2/AM as detailed previously [28,121]. To obtain absolute values of [Mg2+] in single VSM cells, a final concentration of 5 uM mag-fura-2 pentapotassium salt containing either 10 mM(max) or 0-mM MgSO4(min) was used for an in vitro calibration. In some experiments, we added WEB2086 or the SMAse inhibitors, as above. The calibration solutions also contained (in mM ): KCl 115, NaCl 20, and HEPES 5, buffered with NaOH to pH 7.1 under air, at 370C. From these standard solutions, the maximium and minimum intensities of flourescence were obtained at the 335 and 370 nm wavelengths and the ratio of R335/370 was generated . [Mg2+}i was calculated according to the following equation:
and a Kd of 1.5 mM was used for the mag-fura-2/Mg2+ complex [28,121]. B is the ratio of fluorescence intensity of free mag -fura-2 to Mg-bound mag-fura-2 at 370 nm.
The VSM cells were incubated for 18 to 24 hr in either vehicle(NKRB) with and without PAF or modified NKRB containing various [Mg2+]0 with and without PAF , harvested , and assayed for the p65 subunit of NF-kB [81,82,88,97]. In some experiments, we added the PAF inhibitor, WEB2086 (5uM) to the incubation media. The NF-kB p65 (A) rabbit polyclonal IgG immunoaffinity column, purchased from Santa Cruz Biotechnology(Santa Cruz, CA) was used to concentrate the transcription factor [81,82,88,97]. After elution, gel electrophoresis, transfer to a nitrocellulose membrane, and Western blotting, the p65 subunit was assayed using the same IgG for immunodetection [81, 82,88,97].
Primary cultures of rat aortic, piglet coronary and canine cerebral VSM cells were incubated with either vehicle (NKRB) with and without PAF or modified NKRB containing the various concentrations of [Mg2+]0 with or without PAF for the varying intervals of time. In some experiments, we added the PAF inhibitor, WEB2086 (5 uM) to the incubation media. Cells were then lysed, and total cellular RNA extracted using the lithium-urea method [26, 97]. Northern blot analyses were performed as described previously [26, 97]. Blots were hybridized to a randomly, primed specific cDNA probe for c-fos or c-jun and then exposed to Kodak Y-Omat film (Rochester, NY) for 2 to 5 days at -70°C[26,97]. Relative amounts of c-fos and c-jun expression were determined with a densitometric scanner as previously [26, 97].
Cellular levels of pulsed-labeled ceramide were measured utilizing methods we have published previously [81, 82]. Briefly, the VSM cells, in primary cultures, were exposed to vehicle(with or without PAF) or to low [Mg2+]0 (with or without PAF) and [3H] palmitic acid(4-20 uCi/ml) at 37°C, rinsed with fresh NKRB solution or low-Mg NKRB solution and transferred to their respective NKRB’s(under 95% oxygen-5% CO2) for 3 hr. The cells were washed for 5-10 min with unlabeled media, transferred to culture media containing either 0.3 or 1.2 mmol/l [Mg2+]0 and either vehicle(NKRB) or the PAF inhibitor,WEB2086(5 uM). Lipid extractions were performed for phospholipid analysis using techniques similar to those we have reported previously [81, 82,89,90]. Total ceramide cellular levels were determined similar to methods we have used recently employing conversion of ceramide into ceramide-1[32P] phosphate by Escherichica coli DAG kinase followed by lipid separation on high-performance TLC plates [81, 82]. After autoradiography, spots corresponding to ceramide-1-phosphate were carefully scraped into vials, and the radioactivity counted in a scintillation counter (LS-6500; Beckman, CA). Quantitation of ceramide levels and results as picomoles per 106 cells were determined. The results were expressed as umol/ug protein using a Bradford reagent for total protein (Bio0Rad Labs, Hercules, CA) following the manufacturer’s protocol using a 96-well plate[81,82].
Cellular levels of pulsed-labeled ceramide were measured utilizing methods we have published previously [81, 82]. Briefly, the VSM cells, in primary cultures, were exposed to vehicle(with or without PAF) or to low [Mg2+]0 (with or without PAF) and [3H] palmitic acid(4-20 uCi/ml) at 37°C, rinsed with fresh NKRB solution or low-Mg NKRB solution and transferred to their respective NKRB’s(under 95% oxygen-5% CO2) for 3 hr. The cells were washed for 5-10 min with unlabeled media, transferred to culture media containing either 0.3 or 1.2 mmol/l [Mg2+]0 and either vehicle(NKRB) or the PAF inhibitor,WEB2086(5 uM). Lipid extractions were performed for phospholipid analysis using techniques similar to those we have reported previously [81, 82,89,90]. Total ceramide cellular levels were determined similar to methods we have used recently employing conversion of ceramide into ceramide-1[32P] phosphate by Escherichica coli DAG kinase followed by lipid separation on high-performance TLC plates [81, 82]. After autoradiography, spots corresponding to ceramide-1-phosphate were carefully scraped into vials, and the radioactivity counted in a scintillation counter (LS-6500; Beckman, CA). Quantitation of ceramide levels and results as picomoles per 106 cells were determined. The results were expressed as umol/ug protein using a Bradford reagent for total protein (Bio0Rad Labs, Hercules, CA) following the manufacturer’s protocol using a 96-well plate[81,82].
DNA synthesis was examined by incorporation of the thymidine analog bromodeoxyuridine(BrdU) into VSM cells [26]. Subconfluent cells, on 12-mm coverslips, that became quiescent after transfer to serum-free media (DMEM + 0.1% bovine serum albumin[BSA] for 24 hr were reincubated with DMEM + 10% FCS in various concentrations of [Mg2+]0 ( i.e., 1.2, 0.6, 0.48, and 0.3) with or without PAF (with or without 5 uM WEB2086) for 24 hr. At the end of the low Mg2+ with or without PAF (and WEB2086) or the SMAse inhibitors, BrdU (15 umol/l) was added, and the cells were visualized using a BrdU monoclonal antibody( Zymed; San Francisco, CA) followed by the ABC method[26]. The BrdU-positive cell nuclei were counted in four to six high-power fields and related to the total cell number/ high-power field [26].
The methodology outlined by Travers et al [122] and Alapart [123], employing gas chromatography-mass spectrometry (with some modifications for our VSM cells) , was utilized to measure PAF. Lipids were isolated and subjected to GC-mass spectrometry. All measurements were made in triplicate.
Measurement of 8-OH-dg is thought by many experts to be a good method for assessing DNA damage in a variety of cell types [124-126]. Use of liquid chromatography-mass spectrometry is considered the method of choice by most experts [124-126], and is the method employed in our studies [125].
For these measurements, we followed the methodology outlined by Hill et al for cultured VSM cells [127]. As they indicated, we utilized radiolabeled 4-HNE(i.e., [3H]4-HNE, 50 uM, 2.7-3.0 x 106 cpm per well) was added in the presence of low Mg with or without WEB2086. Radioactivity( in the supernatants) was utilized for HPLC analysis. Protein-bound 4-HNE was measured by scintillation counting and normalized as per Hill et al [127].
Fura-2/AM, mag-fura-2/AM, FCS, pluronic F-127, platelet activating factor(PAF), WEB2086, desipramine, scyphostatin, nifedipine, and streptomycin were purchased from Sigma Aldrich Chemical Company(St.Louis, MO). All chemicals used to make up NKRB or modified NKRB solutions were of the highest grades available and purchased from Fisher Scientific Co.(Springfield ,NJ).
Where appropriate, results were expressed as means +/- SEM and were examined for statistical significance by means of Student’s t-tests and analysis of variance (ANOVA) using Scheffe’s contrast test. Values of P<0.05 were considered to be statistically significant.
Results
Table 1 indicates that reduction in [Mg2+]0 results in a rapid, concentration- dependent increase in free [Ca2+]I in rat aortic, piglet coronary and cerebral VSM cells, which plateaus within 2-5 min, similar to that reported previously[26,28,30,51,55,56,72,96]. Although not shown, these elevations in free, ionized cellular Ca2+ were maintained for at least 8-12 hr. The data presented in Table 1 also indicate that when all of the extracellular Ca2+ in the NKRB solution is removed, the basal levels of [Ca2+]I are lowered dramatically 92-98 %, similar to that reported previously. Prior incubation of the VSM cells (30-60 min) with the calcium channel blocker, nifedipine (10-6 M), reduced the low [Mg2+]0 –induced rises in [Ca2+]I by 80-85 % (Table 1). Interestingly, prior incubation of the VSM cells with WEB2086 reduced the low [Mg2+ ]0 -induced rises in [Ca2+]i by 55-65% (Table 2).
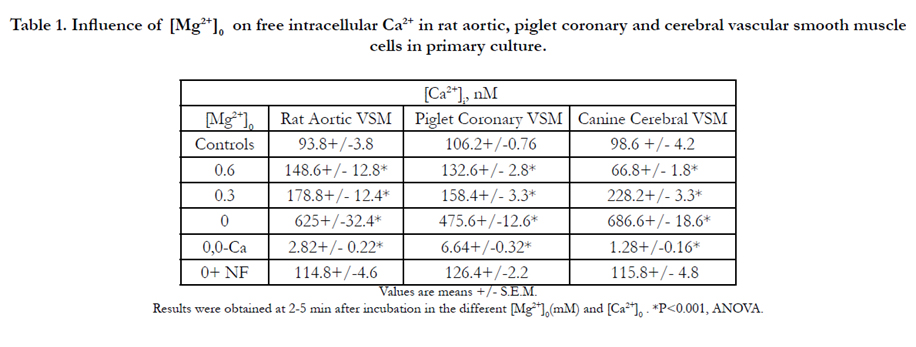

Table 1. Influence of [Mg2+]0 on free intracellular Ca2+ in rat aortic, piglet coronary and cerebral vascular smooth muscle cells in primary culture.
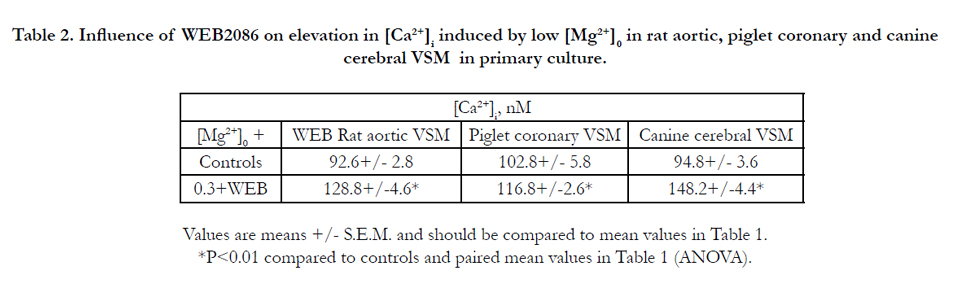
Table 2. Influence of WEB2086 on elevation in [Ca2+]i induced by low [Mg2+]0 in rat aortic, piglet coronary and canine cerebral VSM in primary culture.
Addition of PAF (1-1,000 nM) to the primary cultured VSM cells resulted in a rapid, concentration-dependent rise in intracellular free Ca2+. These elevations induced by PAF were initiated at ~45-60 sec after contact with the VSM cells and peaked at about 2-10 min rising 4- 20-fold over normal, depending on time and concentration (Table 3). If we pre-incubated the VSM cells with WEB2086, most of the PAF-induced elevations in Ca2+ were completely inhibited(Table 3), indicating that the changes in [Ca2+]i were brought about by PAF acting on their specific VSM cell membrane- receptors.
Turning to actions of PAF on intracellular Mg2+, we found that addition of PAF (10-1,000 nM) to the aortic and cerebral VSM cells significantly lowered the [Mg2+]I 10-22 % , depending on concentration, peaking within 3-20 min depending on PAF concentration (Table 4). Concomitant with these changes in cellular free Mg2+ , we noted rises in [Ca2+]I (Table 3). The reductions in [Mg2+]i were somewhat curtailed when the cells were pretreated with WEB2086 (Table 4). Although not shown, similar results were noted in the piglet coronary arterial VSM cells.
Previously, we have demonstrated that exposure of VSM cells to low [Mg2+]0 or feeding animals low dietary Mg intake result in increased levels of both c-fos and c-jun[26,97]. The VSM cells showed upregulation of the proto-oncogenes within 30-120 min after reduction in [Mg2+]0 [26,97]. Addition of PAF (1-1000 nM) to either aortic, piglet, or cerebral VSM cells in NKRB solution also induced formation of both c-fos and c-jun(Fig. 1).
Interestingly, if we preincubated the VSM cells, exposed to low Mg2+ , with the PAF inhibitor(WEB2086), the induction of the proto-oncogenes was attenuated dramatically(see Fig. 2), while the induction of both c-fos and c-jun observed after incubation with PAF(incubated in WEB2086 - NKRB) was almost completely inhibited(Fig. 3).
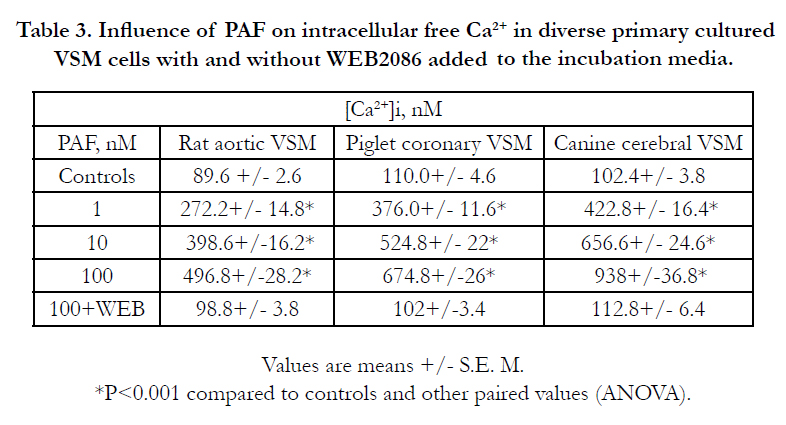
Table 3. Influence of PAF on intracellular free Ca2+ in diverse primary cultured VSM cells with and without WEB2086 added to the incubation media.
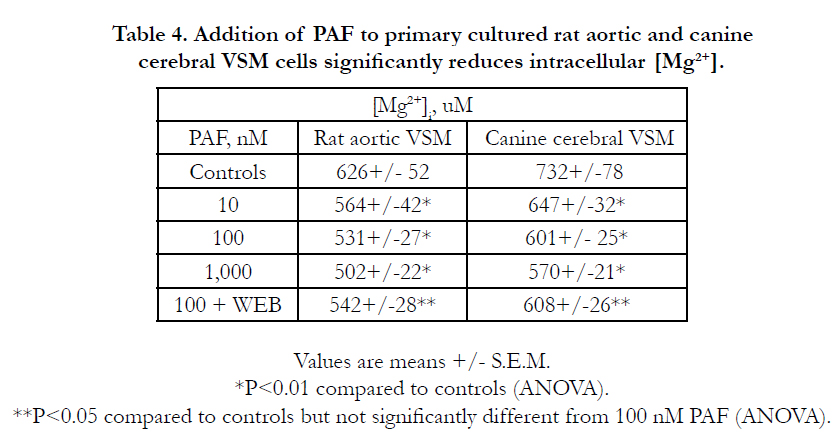
Table 4. Addition of PAF to primary cultured rat aortic and canine cerebral VSM cells significantly reduces intracellular [Mg2+].
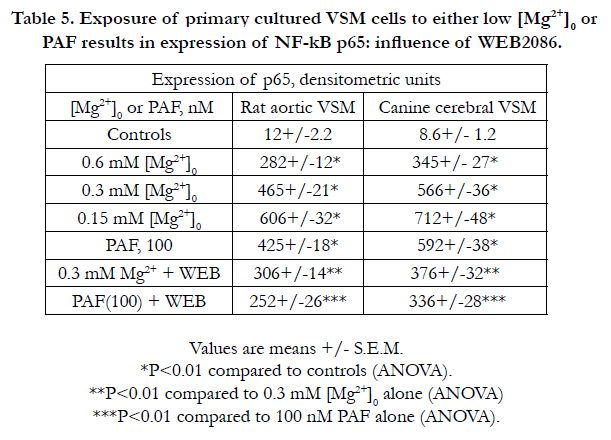
Table 5. Exposure of primary cultured VSM cells to either low [Mg2+]0 or PAF results in expression of NF-kB p65: influence of WEB2086.
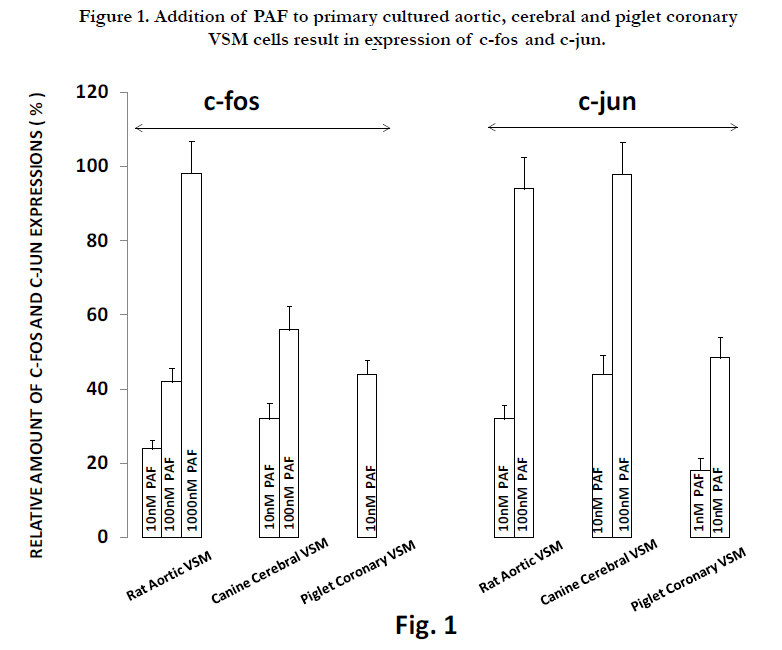
Figure 1. Addition of PAF to primary cultured aortic, cerebral and piglet coronary VSM cells result in expression of c-fos and c-jun
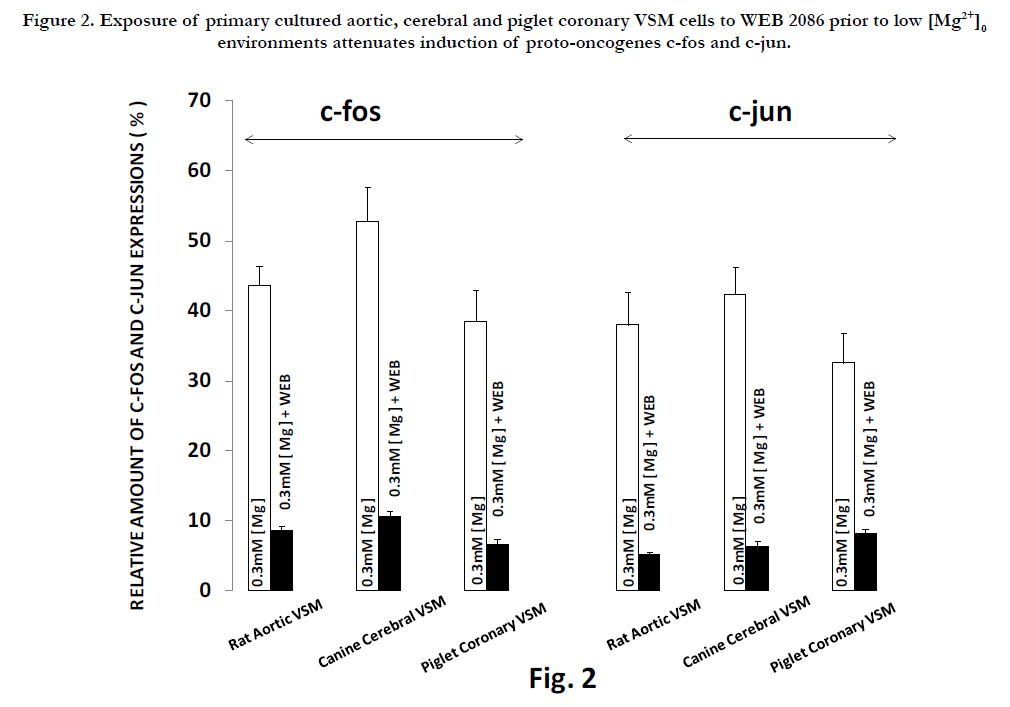
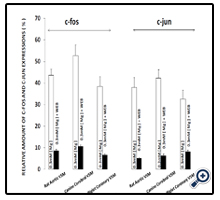
Figure 2. Exposure of primary cultured aortic, cerebral and piglet coronary VSM cells to WEB 2086 prior to low [Mg2+]0 environments attenuates induction of proto-oncogenes c-fos and c-jun.
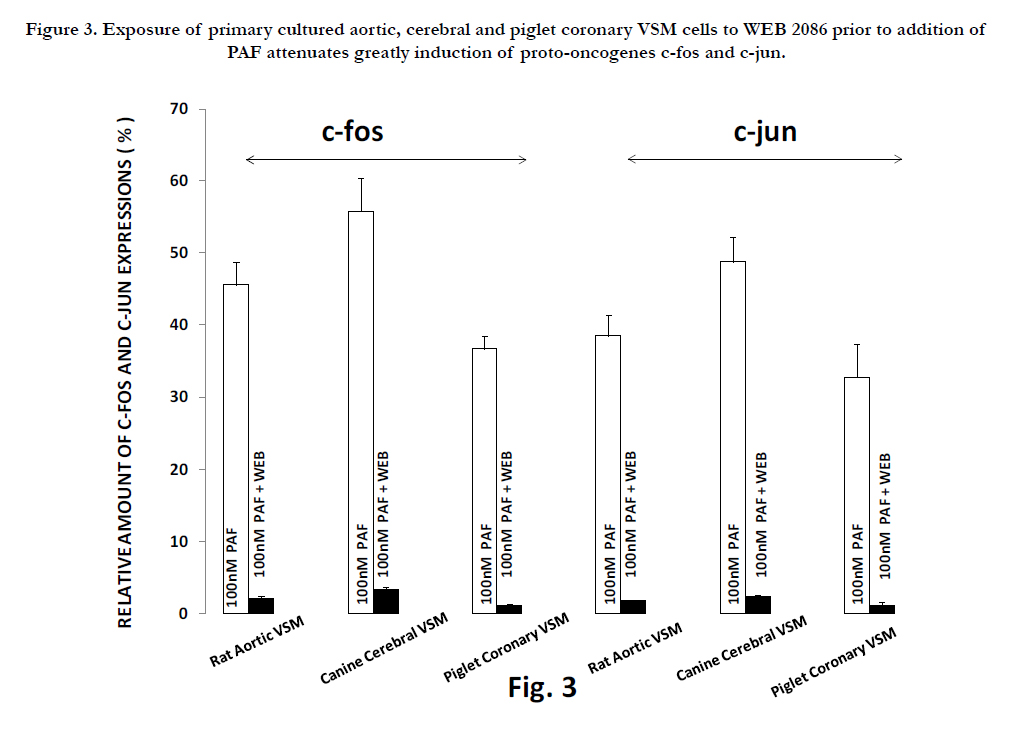
Figure 3. Exposure of primary cultured aortic, cerebral and piglet coronary VSM cells to WEB 2086 prior to addition of PAF attenuates greatly induction of proto-oncogenes c-fos and c-jun.
The results presented in Tables 6 and 7 suggest that exposure of cerebral and aortic VSM cells to either low [Mg2+]0 or PAF appears to induce a concentration-dependent increase in DNA synthesis whereas exposure to either low [Mg2+]o or PAF results, paradoxically, in 6-10 fold increased formation of 8-OH-dg suggesting increased oxidation of the DNA. Incubation of the MgD cells with WEB2086 resulted in decreased synthesis of DNA and only 2-3 fold increase in 8-OH-dg. Although not shown, similar results were noted for the piglet coronary arterial muscle cells.
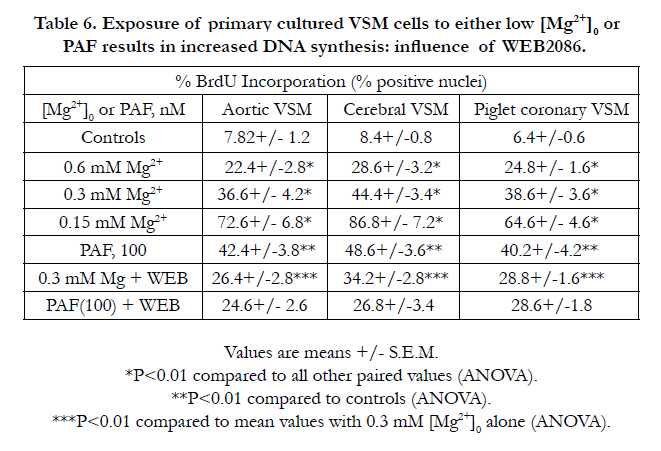
Table 6. Exposure of primary cultured VSM cells to either low [Mg2+n]0 or PAF results in increased DNA synthesis: influence of WEB2086.
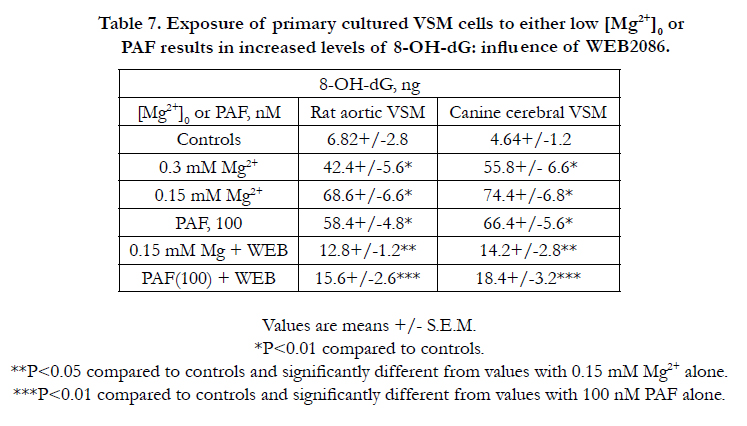
Table 7. Exposure of primary cultured VSM cells to either low [Mg2+]0 or PAF results in increased levels of 8-OH-dG: influence of WEB2086.
Incubation of the VSM cells in low [Mg2+]0 resulted in rises in ceramide, similar to that shown previously [81,82,89,90,95]. Likewise, exposure of the VSM cells to PAF (in NKRB solution) also resulted in rises in ceramide ,but less than that seen in low [Mg2+]0 . Preincubation of the VSM cells in either low [Mg2+]0 or NKRB containing the PAF membrane-receptor antagonist attenuated the rises in ceramide(Table 8). Preincubation of the VSM cells with desipramine and scyphostatin (but not either alone) resulted in almost complete inhibition of the rises in ceramide observed in low [Mg2+]0 and PAF addition (Table 9).
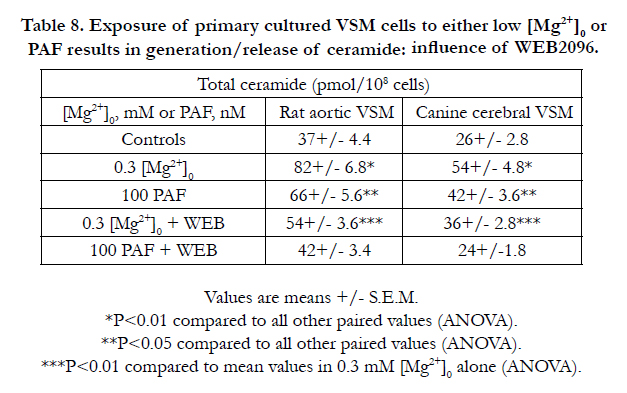
Table 8. Exposure of primary cultured VSM cells to either low [Mg2+]0 or PAF results in generation/release of ceramide: influence of WEB2096.

Table 9. Influence of an N-SMase (scyphostatin) and an acid SMase (desipramine) inhibitor on rises in ceramide levels induced by low Mg2+ and PAF in primary cultured VSM cells.
Incubation of the VSM cells in low [Mg2+]0 resulted in rises in ceramide, similar to that shown previously [81,82,89,90,95]. Likewise, exposure of the VSM cells to PAF (in NKRB solution) also resulted in rises in ceramide ,but less than that seen in low [Mg2+]0 . Preincubation of the VSM cells in either low [Mg2+]0 or NKRB containing the PAF membrane-receptor antagonist attenuated the rises in ceramide(Table 8). Preincubation of the VSM cells with desipramine and scyphostatin (but not either alone) resulted in almost complete inhibition of the rises in ceramide observed in low [Mg2+]0 and PAF addition (Table 9).
The data shown in Table 10 indicate that both PAF and low Mg2+ cause profound increased levels of 4-HNE in aortic and cerebral VSM cells, whereas the presence of WEB2086 inhibits most of the elevations in 4-HNE. Although not shown, similar observations were noted in neonatal coronary VSM cells.
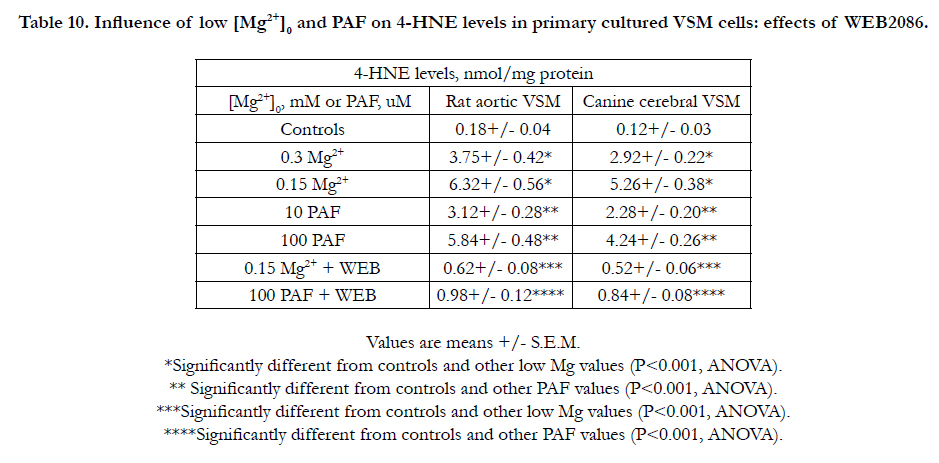
Table 10. Influence of low [Mg2+]0 and PAF on 4-HNE levels in primary cultured VSM cells: effects of WEB2086.
Discussion
To our knowledge, the results reported, herein, are the first demonstration that low concentrations of PAF in at least three different types of VSM cells (from three different mammals), including neonates, results in formation of ceramide, activation of NF-kB, expression of two different proto-oncogenes , and DNA synthesis coupled to rises in intracellular free Ca2+ and significant, albeit small lowering of intracellular free Mg2+ . We also demonstrate that these diverse actions of PAF occur at specific VSM cell PAF membrane-receptors. In addition, we demonstrate , concomitantly, that PAF can induce oxidation of DNA, which also occurs via distinct PAF membrane-receptors. Moreover, we provide new evidence for lipid peroxidation and most likely a pathway for formation of H2O2 in the presence of PAF and low [Mg2+]0 . Lastly, we provide evidence for the idea that many of the VSM cell effects of low [Mg2+]o may be a consequence of synthesis/release of PAF and PAF-like lipids acting via sphingolipid pathways , particularly ceramides. It is important to point out, here, that these are the very first findings to implicate low Mg and PAF in regulation ( and their existence in neonatal coronary VSM cells) of several ceramide pathways in very young mammals, which could have major implications for the numerous, unexplained sudden cardiac deaths , around the world, in infants and young adults [128-134].
The molecular pathways via which ceramides are synthesized are known to be conserved between yeast and mammals [135]. The de novo synthesis of ceramide is brought about via the action of serine palmitoyl-CoA transferase (SPT), 3-ketosphinganine reductase , ceramide synthase (CS), dihydroceramide desaturase and SM synthase (SMS). This reaction directly affects sphingomyelin (SM) , phosphatidylcholine (PC) and ceramide as well as diacylglycerol (DAG) levels. Three of us have previously noted , using primary cerebral and peripheral VSM cells in culture, that a variation in [Mg2+ ]0 influences the cellular levels of SM, PC, DAG, and ceramide [25, 81,82,89,90,93-95,97-99]. Ceramide, either released as a consequence of SMase acting on SM and/or activation of either SPT or SMS or activation of CS is now thought to play important roles in fundamental pathological processes such as cell proliferation, angiogenesis, cell adhesion, diverse immune functions, microcirculatory functions, membrane-receptor functions, programmed cell death(i.e., apoptosis), atherogenesis, inflammation , and aging [135-144]. Over the past two decades, our laboratories have found that low Mg2+ (both in vivo and in vitro) is a fundamental regulator of all of the above major enzymatic pathways and causes upregulation of ceramide in multiple cardiovascular tissues and cells [25,81,82,89,90, 93-95,97-99,150]. Interestingly, we very recently have found evidence for the upregulation of the unique "salvage pathway" for additional synthesis of ceramide in the presence of low [Mg2+]0 in diverse cardiovascular tissues and cells [82]. Although the activation of N-SMase , acid-SMase, SPT-1, SPT-2 as well as SMS by low [Mg2+]0 results in (and ensures) ceramide production in cardiovascular tissues [25,81,82,89,90,93,94,97-99], the activation of CS (by low [Mg2+]0 ) via the "salvage pathway" could result in additional levels of de novo ceramide along with elevated levels of sphingosine [82]. Our recent studies provide new evidence for the existence of the latter ("salvage") pathway in cardiovascular tissues via activation of several protein kinase C (PKC) isozymes, with PKC -zeta playing a dominant role [82]. Previously, we suggested that new therapeutic and prophylactic approaches in the management of some cardiovascular diseases (e.g., hypertension, coronary ischemic syndromes, and atherosclerosis) could be helped/ameliorated with more insight into this Mg-regulated pathway [82]. In view of the present work on PAF, we believe this may be critical in management of inflammatory diseases and atherosclerosis as well as the aging process, and in prevention of sudden cardiac death, particularly in infants and young adults.
Research on PAF and PAF-like lipids has generated considerable interest over the past decade [110- 115, 143-149,155].This research has resulted in the acquisition of a great deal of knowledge into the biology, chemistry, physiology, and pathology of these unique molecules, e.g., PAF synthesis, transport, molecular structures, heterogeneity of PAF receptors, regulation of gene expression by PAF, signal-transducing roles of PAF and PAF-like lipids ( e.g., activation of tyrosine kinases, MAP kinases, and NF-kB), PAF functions in circulatory and immune physiology, reproduction, tumor growth, and infectious diseases.
PAF receptor (PAF-R) signaling is known to be linked to various second messenger networks through phospholipases leading to Ca2+ fluxes. We show in the present study that addition of low concentrations of PAF to primary VSM cells results in rapid rises of intracellular free Ca2+ concomitant with release/synthesis of ceramides via N- and acid-SMAses leading to activation of NF-kB and proto-oncogenes followed by both synthesis and oxidation of DNA, particularly in the presence of low [Mg2+ ]0 . Our new findings indicate that these PAF actions must perforce occur via specific PAF-Rs, as the specific PAF-R membrane antagonist,WEB2086, either prevents or ameliorates these VSM actions of PAF. Our new results also demonstrate that these actions of PAF appear to be dependent on its ability to elevate intracellular free Ca2+ coupled to reduction in intracellular free Mg2+. These new latter observations are the first of their kind, to our knowledge, in any cell type.
The present observations demonstrate that ceramide release (via activation of both N- and acid SMAses) probably occurs as a potential consequence of PAF ( and/or PAF -like lipids) action in VSM cells exposed to low [Mg2+]0. We believe that these observations have a direct relationship to facts that low dietary Mg levels lead to atherogenesis [21,22,25,29], inflammatory conditions [37,38,60-62,67,74,76,77-80], and reduced telomerase activities in cardiovascular tissues and cells[99]; the latter a potential critical factor in the aging process [10-104]. If our present findings, taken together with our previous studies [15,25,26,81,82,89,90-99,107,150], are confirmed they could lead to new approaches to therapy and prophylaxis of cardiovascular and inflammatory diseases with specific antagonists of PAF-R and supplements of diets with Mg, particularly as more than 65 % of The North American population is MgD. In addition, this combination therapy/prophylaxis could lead to longer life spans and better quality of life processes, and probably less sudden cardiac deaths in young adults and infants, provided pregnant women and their offspring are adequately supplemented with Mg in their diets.
We confirm in this report that exposure of diverse VSM cells to low extracellular Mg2+ results in oxidation of DNA [99]. These data, when taken together with other studies [15,26,88-90,95, 152], support the hypothesis that MgD, unless checked early-on, "probably will contribute in a major way to the atherosclerotic and aging processes [15,21,22,25,67,99,106,150,151]". The fact that this event is linked to activation of several major enzymatic sphingolipid- ceramide pathways and PAF production in primary VSM cells provides further evidence for important roles for generation of ceramide and PAF in atherogenesis , inflammation, hypertension , and heart failure; all of these being involved in diverse and major ways in the aging process [99,107,110-115]. More than 10 years ago, working with primary cultured cerebral VSM cells we first demonstrated low [Mg2+ ]0 levels result in rapid , increased formation of molondialdehyde[MDA;]. This Formation of MDA was also noted in diverse cardiovascular tissues extracted from rats subjected to 21 days of MgD [150]. Molondialdehyde is a genotoxic compound that forms a propano adduct with 2'-deoxyguanosine rather than an eteno adduct, which can produce mutations of DNA.
The increased formation of 8-OH-dg observed, herein, and previously in a variety of cardiovascular tissues and cells [124-126,152], which indicates oxidation of DNA, supports suggestions made years ago, that MgD could lead to multiple mutations in the genomes of multiple cell types[153,154]. This could help to also explain what appears to be, seemingly, paradoxical: i.e., MgD-induced DNA synthesis with oxidation of DNA. Alterations in the DNA molecules in MgD could mask oxidation of some DNA and, possibly, still yield a synthesis of DNA as reported here and elsewhere [26]. We have demonstrated , previously, that 21 days of short-term MgD in rats resulted in DNA fragmentation and lipid peroxidation in intact cardiac and VSM cells [150], whereas incubation of cerebral and peripheral VSM cells( in primary culture) resulted in significant synthesis of DNA after exposure to low [Mg2+]0 for a period of 24hr [26]. How can these and the present results be reconciled?
Although all cells in the body (except for red blood cells) continue to develop strategies to preserve the integrity of DNA structure, numerous factors can alter the structure of DNA; such as reactive oxygen and nitrogen species, numerous extrinsic chemical molecules, and UV irradiation to name a few [124-126, 156]. Each day, DNA of diverse mammalian tissues and cells (including cardiovascular components) receive multiple assaults from , for example, guanine methylations, cytosine deaminations, spontaneous depurinations, single- strand breaks, double-strand breaks, and oxidative lesions, among others[87,110,116,119,124, 152,153,156,157]. Every one of these insults induces source-specific damage of DNA. For example, external chemical molecules can cause alkylation of DNA by methylnitrosourea [152,158]. Cancer therapeutic agents such as mitomycin C and cisplatin can induce cross-links in DNA [152,158]. Reactive oxygen and nitrogen species can induce very-specific base modifications in the DNA [87,110,116,119,124,152,153,156-158]. UV light is well-known to induce strand breaks in DNA, DNA-DNA cross links, and DNA-protein link formations [158]. Interestingly, each one of these assault species cause different DNA aberrations leading to different DNA repair processes [152,153,156-158]. All of these diverse situations would thus not only alter DNA in different ways, affect gene regulation via diverse transcription factors, but modulate DNA damage/ repair processes. A few years ago, using a short-term model of MgD , we found that cardiovascular tissues and cells demonstrated marked reductions in glutathione content coupled to activation of both e-NOS and n-NOS [95]. Since eNOS activity has been reported recently to be important in activation of PAF in endothelial cells [114,115], our findings of low Mg2+ -induced synthesis/generation of eNOS in cardiovascular tissues and cells may trigger (and be linked), in part, to activation of the SM-ceramide pathways [95], DNA damage and synthesis.
Interestingly, it has been reported recently that DNA damage and the DNA damage response have been identified in human atherosclerosis [159, 160]. As we demonstrated approximately 25 years ago, in a rabbit model, suboptimal dietary intake of Mg (similar to that which 65% of the North Americans ingest daily) results in rapid atherosclerosis with plaques over more than 60% of the aortic and coronary arterial surfaces [21]. Recent human studies, using serial angiography coupled with postmortem studies, suggest that many plaques appear to invade coronary arterial walls months before myocardial infarctions and in the absence of blood clots [161]. Our previous studies indicated that low [Mg2+ ]0 environments lead to coronary vasospasm and ischemia prior to plaque formation[3,25,29,53,120,151,162]. Depending on the diverse type of DNA modification, cells would exact different repair processes in situ in order to remove such damage. In contrast to the in vitro situation , cells in primary culture ( under tightly controlled environments, such as the present study) would be expected to show possibly different effects , e.g., with low [Mg2+ ]0 ,i.e., DNA synthesis (over short incubation times) as shown here and elsewhere [26]. Thus, it is important to keep in mind that both DNA damage and synthesis could be expected in diverse tissues and cells in vivo, depending upon time and circulating/ cellular levels of free ionized Mg. From this argument, when taken together with data presented herein and elsewhere [3,4,6-9,15-31,34,37-39,41,48-56,59,65-67,74,76-82,88-99,105-107,150,151], low Mg environments could be expected to exert multiple effects/ actions in atherogenesis, inflammatory conditions, on the aging process, and thus on cardiac stability.
Approximately eight-10 years ago, it was shown by Lee et al. in HL-60 cells and Xenopus oocytes that a membrane-associated, CoA- independent transacetylase can transfer the acetate group from PAF to sphingosine and form ceramide [163]. Whether or not this enzyme-pathway exists in VSM cells and cardiac muscle cells is not known. However, since we have recently demonstrated that low [Mg2+ ]0 can result in formation of some sphingosine and ceramide via the "salvage pathway" [98], we are tempted to speculate that the pathway discovered by Lee et al leading to PAF and ceramide may be operative and important in the cardiovascular effects of MgD. Almost 20 years ago, we reported, using the same VSM cell(herein) and proton-NMR spectroscopy that low Mg2+ environments resulted in rapid formation of PAF, PAF -like lipids and plasmalogens[89]. Lee's work suggests that a number of PAF-like lipids and plasmalogens could be transacetylated (as above) to form a variety of important lipids that could play major "roles in modifying the biological responses of PAF by generating a variety of its analogs" [110]. These possibilities, obviously, remain to be investigated and could be quite important in atherogenesis, inflammationm the aging process, and cardiac stability.
Over the past two decades, a considerable amount of knowledge and insight into the regulation of PAF-Rs has accumulated [144-149]. But, despite these efforts, specific regulatory systems for the PAF-Rs, in diverse tissues and cells, remains somewhat nebulous. It is currently thought that any factor that can affect binding of PAF (and PAF-like lipids) to their membrane-receptors or to subsequent PAF-dependent-signaling events would be likely candidates as regulators of specific PAF-Rs [111-114, 147-149]. Our present work could be suggestive of a role for Mg2+ in regulation of PAF-Rs in VSM cell membranes. More than 50 years ago, our labs and other workers [5, 49, 50, 53, 164-166] reported that Mg2+ ions are needed for the binding of certain neurohumoral molecules (e.g., vasopressin, oxytocin, catecholamines, angiotensin II) to their receptors on VSM cells as well as their contractile actions. Although not shown, we have found that [Mg2+]0 also appears to be important in the VSM contractile actions of PAF [ unpublished studies]. So, it would appear that from the present studies and others [3-5,8, 14, 15-32 34, 35, 38,40,48-56, 59,65, 72-74,81, 88-100, 105-108] that Mg2+ plays important and diverse, multiple regulatory actions in VSM cells. Much more investigation will, however, be required to reveal the specific interactions of PAF, PAF-Rs, and VSM cells.
Obviously, much work (including the present studies) has been brought forth to show that PAF is a major mediator of multiple intracellular interactions and pathways. Our work indicates that PAF is a regulator of several downstream, important transcriptional pathways in VSM cells, i.e., NF-kB and proto-oncogenes, and, most likely, sphingolipid-ceramide pathways.
Platelet-activating factor (PAF) is a phospholipid mediator which displays multiple biological and pathophysiological attributes, running the gamut from inflammation to cell differentiation and proliferation. As is well-known, PAF interacts with specific membrane PAF-Rs to initiate all cellular responses via specific G-protein-coupled receptors. Although these initiated membrane-receptor phenomena are mostly established for several tissue-cell systems [111-114,144-149], exactly how PAF and PAF-Rs cause vascular remodeling in hypertension and atherosclerosis is not clear. A little more than 15 years ago, PAF was identified as a molecule that stimulated activation of NF-kB [113,114,148].
NF-kB is now known to be a prime regulator of growth processes, differentiation, cell migration, and cell death [for reviews, see ref. 83-86], all factors required for vascular remodeling in hypertension and atherogenesis [1,87]. NF-kB is clearly a major transcription factor and a pleiotrophic regulator of numerous genes involved in inflammatory processes and epigenetic phenomena [83-86]. NF-kB is now thought to be a pivotal transcription molecule involved in atherogenesis , hypertension and stroke [1,15,26,29,81,87,88,97,164,165,168]. As of now, it is still not clear as to what factor (s) initiates expression of these molecular events. We were the first laboratory to suggest and provide evidence for a role for activation of NF-kB in development of low [Mg2+]0 -induced atherosclerosis [26]. We demonstrated that when VSM cells were exposed to low concentrations of [Mg2+]0 , a concentration-dependent upregulation of NF-kB took place; the lower the [Mg2+]0 , the faster and greater the upregulation of NF-kB [26,81,82]. The present results confirm and extend these findings, as do other subsequent studies in endothelial cells [167]. However, herein, we demonstrate that lowering extracellular Mg2+ also results in a rapid rise in active PAF prior to NF-kB activation. Simultaneously and concomitant with the latter we have shown that two proto-oncogenes, c-fos and c-jun, are formed, most likely as a consequence of PAF synthesis and concomitant release / synthesis of ceramide . The fact that the specific PAF-R blocker, WEB2086, inhibits the PAF-stimulated formation of the proto-oncogenes in the VSM cells in our present experiments lends considerable support for this contention.
Several lines of evidence suggest that ceramides are important in cytokine generation and in cytokine-induced apoptosis [81, 82,140, 167,168], events which are pivotal in vascular remodeling in atherogenesis and hypertension [1, 13]. In this context, we have shown both in short-term MgD in intact rats and in vitro, using primary cell culture of VSM cells, exposed to low extracellular Mg2+, that these pertubations result in synthesis/release of at least 12 different cytokines and chemokines [81,82], known to be involved in development of atherosclerosis and hypertension . It is important, here, to point out, again, that phenotypic transformation of VSM cells go from a differentiated state to a proliferative one during vascular injury and inflammation, early events in atherogenesis [1, 13,109]. Cytokines, in particular, such as TNF-alpha, induce VSM migration and proliferation mediated by several molecular pathways such as ERK and p38MAPK [155,164-166, 168] , which we have found to be upregulated in VSM cells exposed to low [Mg2+ ]0 [54,56]. It is of considerable interest to note, here, that these 12 different cytokines and chemokines (which we have shown to be upregulated in MgD) have also been reported to be generated in the presence of PAF [113-115,146]. Moreover, cytokines such as TNF-alpha, stimulate formation of superoxide dismutase, reactive oxygen and nitrogen species, and hydrogen peroxide [154], all of which we have found to be upregulated in MgD in cardiovascular cells and tissues [15,25,26,81,82,88,89,90,91,95,151, present studies with HNE]. We, thus, believe that MgD and its stimulation of PAF present a powerful combination with the generation/release of ceramide for a prime mechanism for vascular remodeling of arteries and arterioles which perforce are a sine-qua-non in atherosclerosis and hypertension. The fact that the PAF-R blocker, WEB2086, inhibits a great deal of NF-kB activation, proto-oncogene formation, and lipid peroxidation (seen in present work with inhibition of 4-HNE) in VSM cells, exposed to low [Mg2+]0, lends considerable credence to our hypothesis. It is of considerable interest, here, to note that in our rabbit model of MgD-induced atherosclerosis, there were early signs of histochemical identification of PAF in the lipid - laden arterial VSM cells [unpublished findings].
Over the past 20 years, data has accumulated, from many sources, that both c-fos and c-jun proteins (components of AP-1 transcription factor) are critically involved in the mediation of nuclear events provoked by numerous stimuli [83-87, 172-174]. The proto-oncogenes c-fos and c-jun are known to participate in numerous pathophysiological processes including signal transduction, cell growth, differentiation, and cell transformation [83-87]. Both c-fos and c-jun are thought to play roles as potent inducers of apoptosis in several cell types [83-87]. The proto-oncogenes c-fos and c-jun encode nuclear phosphoproteins. Members of the c-fos and c-jun families dimerize to preferentially bind AP-1 sites with very high affinity. We believe the upregulation of the proto-oncogenes , in the present study , by PAF and low [Mg2+]0 , as well as our previous experiments in vivo and in vitro , support the notion that c-fos and c-jun upregulate genes controlling cell growth , differentiation, and cell transformation via AP-1 binding genes, all of which are essential in atherogenesis and hypertension. In normal VSM and cardiac muscle cells, the expression of growth factors and their receptors is carefully controlled. However, over-activity of the c-fos and c-jun proto-oncogenes could result in unregulated cell proliferation like that observed in atherogenesis and hypertension. Our new findings support the hypothesis we advanced previously: "that overexpression of c-fos and c-jun in the cardiovascular system of MgD animals and humans may play an important role in the course of tissue inflammation and the inflammatory response observed in the atherogenesis noted in MgD animals" [83-87].
The observations presented, herein, when taken together with other studies from our labs [25,26,29,97], indicate that low [Mg2+]0 is a powerful stimulant for formation of both c-fos and c-jun proteins in primary cultured VSM cells. In addition, it now appears from the present work that PAF, acting via specific PAF-Rs on VSM cells, is also a powerful stimulant of the proto-oncogenes acting probably via ceramides and NF-kB, and most likely PKC activation coupled to Ca2+ entry and intracellular release [26].
Sudden infant death syndrome has become a significant problem without an agreed-upon pathological mechanism(s) [for reviews, see 128-134]. Explanations run the gamut from hypoxia, gene mutations, cardiac conduction abnormalities, inherited channelopathies, unknown infections, diaphragmatic dysfunctions, central hypoventilation, susceptibility to ventricular arrhythmias, etc [128-134]. Pediatric sudden cardiac arrest likewise seems, for the most part, to be growing problem around the world with little in the way of an acceptable pathogenic mechanism(s) [ for reviews, see 130, 131,133,134]. Sudden cardiac death (SCD) accounts for approximately 20% of nearly all deaths in Western countries with multiple possible, but little agreed-upon, explanations . SCD in the young (< 35 yrs of age) has a structural/genetic basis in only about 20% of all cases in the Western world [128-134]. In these SCD cases, there is no structural/pathological evidence for any heart abnormalities on autopsies [128-134].
Approximately 35 years ago, our laboratories reported that mammalian coronary arteries( including from humans) demonstrate intense vasospasm as [Mg2+]0 concentrations are progressive lowered in in-vitro studies; the lower the reduction in [Mg2+]0 , and the smaller the coronary vessel, the more intense the coronary arterial vasospasm [3]. No vasodilator, including Ca2+ channel blockers can alleviate these intense vasospasms [3-5; unpublished findings ]. These contractile actions of low Mg2+ were potentiated in the presence of neurohumoral and circulating vasoconstrictor agents, such as angiotensin II, vasopressin, serotonin, norepinephrine, and a variety of peptides [3-5, 19,49 ,50]. The present studies not only provides further evidence for this concept, but now implicates formation/ synthesis of PAF , most likely, in these unique coronary arterial vasospasms. As we have shown, previously, that piglet coronary arteries behave similarly in low Mg2+ (i.e., intense vasospasm)[120 ], the present findings using primary culture of piglet coronary arterial vessels lead us to conclude that low dietary levels of Mg in pregnant women, infants and children must be taken into consideration as a major underlying mechanism for unexplained SCD in infants , children and young adults.
It is of considerable interest to point out here that several studies have been published which indicate that infants and children in the U.S.A. demonstrate a much higher percentage of abnormally lowered serum total and ionized Mg levels when compared to adults 35-60 years of age[14,58,62,175-178 ]. Moreover, our studies on infants, children and pregnant women have shown a high percentage of abnormally low serum ionized Mg levels as well [ 58,62,64,179-181 ]. Taken together, we believe such clinical studies, collectively, provide substantial evidence for our hypothesis that unexplained SCD in infants and young children, possibly, is due, in large measure to abnormally low serum, VSM , and cardiac myocyte Mg levels. With respect to the latter, we have shown, working with perfused rat hearts , that even short-term MgD results in reductions in a variety of hemodynamic functions, i.e., cardiac output, coronary flows, stroke volume, developed pressures, and cellular high-energy phosphate levels with concomitant cellular Ca2+ overload[107,162 ]. We showed more than 40 years ago that Mg2+ blocks the entry and release of Ca2+ from both VSM and cardiac myocytes [53,107 ]. This together with the adverse effects of low Mg on both hemodynamics and energy production (and utilization) would perforce seriously compromise young hearts and result in SCD. Whether or not PAF plays an intermediate or trigger role in the effects of MgD on the heart's physiological functions remains to be determined.
Aging is clearly agreed to be critical in the etiology of metabolic decline in most human subjects as they near their 65th birthday. A great many human subjects at 65 years of age demonstrate clear signs of metabolic and physiologic functional decline, atherosclerosis in most major arteries, high blood pressure, diverse cardiovascular diseases, and often type 2 diabetes, which contribute in major ways to congestive heart failure by their 75th -85th years. It must be pointed out, here, that all of these aberrations have been shown( or to be associated with) both experimentally and clinically, with the presence of MgD when they have been looked for [14-18,25,26,28,29,31-36,52,57-71,105-107,150,151,167 ]. It is well- known that the aging process is associated with elevations in blood/serum/tissue levels of many of the same proinflammatory cytokines and chemokines found in MgD animals, e.g., IL-1, IL-2, IL-6, TNF-alpha, among many others [for recent review, see chapters in ref. 185]. Reduced levels of telomerases are known to be associated with elevated levels of several cytokines(e.g., TNF-alpha) in a number of aged cell types as well as in serum and cardiovascular tissues and VSM cells of MgD animals [ for review, see 99 ]; these phenomena being associated with( and correlated to ionized Mg levels, ceramide generation, and activation of NF-kB and proto-oncogene pathways) [99 ]. It should be pointed out, here, that normal amounts of telomerases in all cell types are required to promote efficient cell cycle kinetics and normal cell growth[100-103 ]. MgD is well-known to promote disturbances in cell cycle kinetics [186-188 ] via reactive oxygen and nitrogen species [15,25,26,29,33,67,88,91,95,98,99,106,151 ], most likely acting to downregulate telomerases [99,100-103 ].
Recently , we showed, in animals subjected to 21 days of MgD, that telomerase levels are downregulated coupled to oxidation of DNA and increased levels of p53 [99 ]. Such data supports the idea that MgD could lead to multiple mutations in the genomes of multiple cell types . Our previous studies, when viewed in light of the present experiments, would support the hypothesis that mutations and transformations of VSM, endothelial , and cardiac myocte cells caused by ceramides, MgD and oxidation of DNA (seen in atherogenesis, hypertension, stroke, and congestive heart failure) may play major roles in the aging process, thus leading to multiple cardiovascular changes including inflammations of the vascular walls, high blood pressure( due to formed elemental changes, release of ceramides, release of cytokines, excess lipid wall deposition, lipid peroxidation, etc.), cardiac dysfunctions, and eventual cardiac failure and/or strokes.
Almost 30 years ago, two of us reported , for the first time, the importance of the endothelial lining of blood vessels in the responses of arterial muscles to reduced levels of [Mg2+ ]0 [ 180 ]. More recently this work has been expanded by our laboratory and others [167,181 ], reinforcing the roles of Mg in control of the microcirculation, blood pressure, distribution of blood flow, inflammation and atherogenesis[4,14,15,17,18,19,20,25,29,34,49,53,59,68,74,77,78,79 ]. We have found , as stated above, that short-term MgD causes marked reductions in the cardiovascular tissue/cell levels of the free radical scavenger glutathione(which is correlated with reduced Mg levels) coupled to fragmentation of DNA(and oxidation) and activation of diverse nitric oxide synthases [ 15,25,81,82,89,90,91,93-95,97-99,150]. Such data, when viewed in light of the present work, with PAF, supports the hypothesis that MgD would be expected to cause mutations in many types of cells. These aberrations would promote accelerated aging processes in many bodily tissues and cells. In addition, such alterations in cell types and recruitment of a multiplicity of diverse white blood cells are the hallmarks of atherogenesis ,and inflammatory responses needed to initiate and sustain the atherosclerotic process.
Approximately 40 years ago, Dr. Russell Ross and colleagues advanced the hypothesis that atherosclerosis is an inflammatory disease brought about by injury to the endothelial surfaces of blood vessels in the macro-and micro-circulations[see ref 109, for summary of their hypothesis]. The hypothesis stated that different forms of injury will result in numerous dysfunctions in the homeostatic properties of the endothelium, e.g., increase in adhesiveness of leukocytes and/or platelets, alteration in the procoagulant properties , formation/release of cytokines/chemokines and growth factors. Usually, inflammation is defined as a response of microcrculatory blood vessels and the tissues they perfuse to infections and damaged tissues which bring cells and host-defense molecules directly from the circulation to all the diverse sites where they are required, in order to eliminate/degrade the offending agents [for reviews, see 223,224, for reviews]. The mediators of the defense mechanisms include white blood cells, phagocytic leukocytes, antibodies, chemokines and complement proteins[13, 223-226] .Most of these cells and molecules are recruited ,when needed, from the circulating blood itself. The inflammatory process brings these cells and molecules to the damaged or necrotic tissues. The absence of the inflammatory process would allow infections to continue unchecked, prevent wounds from healing, and result in festering sores/wounds. A typical inflammatory response develops in a sequential manner: recognition of the offending agent(s) by host cells and molecules; recruitment of leukocytes and plasma proteins; activation of the leukocytes and plasma proteins to destroy and eliminate all offending substances; control and termination of the reaction(s); and finally repair of the damaged tissue(s). During the normal inflammatory process, leukocytes migrate across the venous capillary walls through the endotheliun due to increases in permeability and move to the site(s) of injury via chemotaxis . The normal mediators for these processes to take place are: adhesion molecules; and cytokines and chemokines. Interestingly, all of these mediators are needed for atherogenesis and are released/formed in MgD [ 14,15,23-27,29,34,38,41,81,82,88-91,95,97-99,167,226 ]. If, however, the inflammatory response is not curtailed, or effectively neutralized, the inflammatory response will go-on and stimulate migration and proliferation of VSM cells which will become intermixed with the inflammatory cells and protein components to initiate and form an intermediate lesion(i.e., beginning of the atherosclerotic process). If these processes go-on unabated , the arterial walls will thicken and initially dilate to compensate, to a point, then undergo a remodeling process in which the normal contractile VSM cells are transformed into a new non-contractile phenotype [109, 223, 224 ] . At every stage of this process macrophages, monocytes, and T-lymphocytes will be attracted and adhere to the arterial-endothelial walls[109,223,224 ]. Activation of these cells results in release of hydrolytic enzymes, cytokines, chemokines, and growth factors, as stated above. These factors will sustain and perpetuate the atherogenic process forming, eventually, fibrous tissue and further enlargement of the lesion, which will overlie a core of lipid resulting in plaques and necrotic tissue. MgD ,itself, results in release of hydrolytic enzymes, cytokines, chemokines, and growth factors involved in the early stages of the atherogenic process demonstrating lesions and plaques on the inner endothelial surfaces [23,25,38,74,81 ,98,167 ]. In developing atherosclerosis, each plaque has a cap that retains cholesterol, and inflammation inside the plaques can dissolve the fibers, but , suddenly, the cap ruptures ,spilling cholesterol into the insides of the arteries which can promptly cause clots that can completely block the flow of blood into the microcirculation of the surrounding tissues. It has been shown by our lab[81,93,94,unpublished findings ] and others that ceramides, in increased levels, are found in the atherogenic process(i.e, mixed in the plaques), at least in rabbits [227,228 ], and most importantly, are stimulated in MgD [25,81,93,94,unpublished findings ].
Using open-and closed window chambers[see ref 229-232, for methods] implanted in the cerebral cortex ,of living rats and mice in situ, on MgD diets for 21 days [A.Gebrewold, A. Carella, B.T. Altura & B.M. Altura ,unpublished findings ], as well as in the skin and skeletal muscles of MgD rats and mice , we found increased numbers of white blood cells(including monocytes, phagocytic leukocytes, and lymphocytes) on the endothelial surfaces of microcirculatory blood vessels using intravital microscopy at high magnifications(e.g., up to 4,000x normal). We have shown recently that after 12-18 weeks on MgD diets, which produces increases in arterial blood pressure[17,18], these microvasculatures demonstrate less rolling of the white blood cells and much more adherence of the white blood cells to the endothelial surfaces[A. Gebrewold & B.M. Altura, unpublished findings ]. Does PAF play any role in these inflammatory events?
PAF is known to play major roles in both inflammatory responses and atherogenesis[111-115 ]. A variety of the circulating blood cells(e.g., leukocytes, platelets, basophils, macrophages, and endothelial cells) can elaborate PAF[113-115 ], including the VSM cells shown herein. In addition, like MgD , PAF produces vasoconstriction of blood vessels and VSM[111,113-115 ] , as do several of the ceramides[15,92,137,233,234 ]. A number of investigators employing intravital microcirculatory techniques, similar to those used by our group, have clearly demonstrated that PAF increased the numbers of white blood cells in the microvessels concomitant with intense vasoconstriction-spasms with increasing dosing of the lipid mediator, less rolling , and increased adherence of the leukocytes to the endothelial surfaces with increases in vascular permeabilty[235-240]. Using open and closed chambers implanted in rodent cerebral cortex and skeletal muscle, we have observed similar phenomena[A. Gebrewold, B.T. Altura & B.M. Altura, unpublished findings ]. In addition, we have reported that a variety of ceramides produce similar microcirculatory actions in rodent cerebral, cutaneous and skeletal muscle circulatory systems, including increased permeability of the postcapillary venular walls, major sites of inflammatory reactions [234; A. Gebrewold et al, unpublished findings]. Collectively , these in vivo microcirculatory findings strongly support the hypothesis that both PAF and ceramides induce similar, true inflammatory responses in diverse vascular beds in diverse mammalian species.
Having the above, large number of in vivo microcirculatory studies, from our labs and others, we hypothesize that since MgD results in most of the attributes of early atherogenesis, including synthesis/release of PAF and ceramides , PAF and ceramides mostly likely are important, if not critical, mediators released/synthesized in cardiovascular tissues to initiate the early atherosclerotic process.
When the current work is viewed in light of an accumulation of other pertinent data, published elsewhere [15, 25,26,81,82,89-91,93-95,97-99,150 ], we conclude that upregulation of N- and acid SMases (probably in large measure by PAF formation and Ca2+ entry/intracellular release), which are induced in all cardiovascular tissues and cells, in at least three different mammalian species [see above refs. ; present studies], in short-term MgD, results in the formation/release of ceramides and multiple signal transduction pathways; e,g., activation of PKCs, activation of MAP kinases, activation of NF-kB, activation of proto-oncogenes, activation of p53, activation of AP-1, release of cytokines/chemokines, as well as other cell signals . As reviewed above, and elsewhere[5,15,19,25-27,29,30,34,49-51,53,72,73,107,120,189-191 ], Mg2+ plays pivotal roles in control of cellular Ca2+ uptake, subcellular content, and subcellular distribution in smooth muscle cells, endothelial cells, and cardiac muscle cells [5,19,26,30,49,50,53,72,92,107,120,162,190,191 ]. Mg2+ regulates cardiac hemodynamics, regulation of the microcirculation, vascular tone, vascular reactivity, endothelial functions, clotting of blood formed elements, carbohydrate , nucleotide, and lipid metabolism, prevention of free radical formation, and stabilization of the genome [4-11,14-23,25-37,49-59,74,75,79,81,82,88-99,105-107,150,151,162,167 ].
With the advent of new Mg2+ - ion selective electrodes, a new era has been ushered -in for the diagnosis and potential treatment of hypertension, ischemic heart disease, atherogenesis, peripheral vascular diseases, preeclamptic- eclamptic disorders in pregnant women, diabetic-related vascular diseases, sickle cell anemic-related vascular diseases, trauma-shock injuries, cardiac failure, strokes, and organ transplantation [15,17,25,27-31,35,57,58,60-70,89-91, 192-208 ]. Since most of these disorders ( or clinical situations) become worsened with age, or take place in the natural aging process, and are known (from mostly our studies) to be associated with varying degrees of MgD, the role of Mg in carbohydrate , lipid, nucleotide, RNA, DNA, and protein metabolism, and the effects of Mg deficiency, particularly on sphingolipid metabolism, appear to play vital roles on telomerase activities and telomerases, and, most likely, the genome.
In 2010, we first suggested that MgD , by itself, probably acts as a genotoxic agent [93,94 ]. One of ceramide's major pathophysiological actions is its ability to induce cell differentiation and cell transformation [136-141 ]. Such actions in MgD could aid in explaining, to a large extent, transformation of contractile VSM cells to a new phenotype in atherogenesis--i.e., VSM cells that no longer contract or relax, but become synthetic machines for a variety of cytokines, chemokines, and growth factors. Abnormal cell differentiation, transformation , and growth are pivotal events in the developmnt of atherogenesis, hypertension, and progressive cardiac failure. Hyperplasia and cardiovascular hypertrophy are common events in aging, hypertension, and progressive cardiac failure [1,2,185 ].But, the precise mechanisms are not completely understood.
PAF, ceramide, and the tumor suppressor factor p53 are now known to play key roles in cell transformations, growth, apoptotic events, and the aging process [83-87,110-115,135-141,145,147-149,209,210 ]. PAF, ceramides and p53 all can induce cell cycle arrest (and senescence), induce programmed cell death, and are associated with DNA damage( i.e., genotoxic events)[83-87,110-115,135-141,145,147-149,209,210 ]. As reviewed above, and elsewhere [15,16,26,29,74,91,99,107,151, ], MgD can produce all of these events in multiple cell types, including cardiac and VSM cells. Over the past 12 years[15,25,81,82,89-91,93-95,97-99,211-218 ] (including studies herein), we have shown that MgD ( via activation of p53[ 94] and now PAF) is a" driver" of ceramide synthesis/release via multiple cellular pathways in diverse cardiac and VSM cells, including, now , neonatal coronary VSM cells.
It should be recalled, here, that atherosclerotic plaques in the vascular walls of hypertensive human subjects demonstrate considerable DNA damage, activation of DNA repair pathways, increased expression of p53, increased expression of proto-oncogenes, activation of NF-kB, oxidation, lipid peroxidation, and apoptosis[110,157-160 ]. We have found all of these pathobiological alterations in MgD experimental animals. We do not believe this is a coincidence. Experimentally, MgD results in accelerated atherosclerotic plaques in rabbit and mice arterial walls [21,22,24 ], which has been associated with increased levels of p53 [94 ] and PAF [110,115 ] in the thickened atherosclerotic plaques and decreased levels of telomerases [unpublished findings]. We, thus, hypothesize that MgD plays key roles in the generation of atherosclerotic plaques , coronary arterial diseases, and hypertension via the upregulation of N-SMases, acid-SMases, p53, ceramide synthase, PAF generation ,and ceramide production as well as downregulation of telomerases, particularly as the majority of individuals who consume Western-type diets have 45-70% short falls in daily Mg intake. All of the foregoing events could be expected to be accelerated in the aging population, particularly in old-age and nursing homes where the daily intake of Mg often approaches deficits near 80-90% [14,67 ].
Various modifications take place daily and over various periods of time via epigenetics [220,221]. Epigenetic modifications are chromatin-based modifications that alter expression of genes without altering the DNA sequence itself [ 220-223 ]. The most common epigenetic modifications include methylation of DNA, posttranslational modification of histone proteins, and RNA-based mechanisms[222,223]. These biochemical and molecular events can modulate the tertiary structure and, therefore, the accessibility of promoter DNAs or encoding transcription factors and numerous regulatory molecules that, in the end, finally alter transcription [220-223 ]. In these ways, epigenetic pathways are pivotal in the regulation of VSM, endothelial, and cardiac myocyte gene expressions . Alterations in gene expression, for example, would change a normal contractile VSM cell into a cell that becomes a synthetic machine for numerous factors needed for atherogenesis, i.e., cytokines, chemokines, growth factors, etc.; the VSM cell would no longer be a contractile functioning cell type under such transformative conditions. We believe prolonged MgD does this and more.
Last, but not least, our results bolster the idea, we have espoused previously, that water intake(e.g., from tap waters, well waters, bottled waters, beverages using tap/well/spring waters, or desalinated waters) in humans should contain at least 25-40 mg Mg2+ /l [81, 82, 91,93-95,97-99,150 ] . The latter inclusion in our diets should go a long- way towards the prevention of cardiovascular diseases and ameliorate the aging process of bodily tissues and cells in humans worldwide. In 2009, The World Health Organization recommended our guidelines of 25-40 mg Mg2+/l to be included in drinking waters for human consumption[241].
Acknowledgements
The authors appreciate the valuable technical assistance of J-P Liu, Kishan D. Patel, and Tara S. Shah. Some of this work was supported, in part, by NIH grants to B.M. Altura and B.T. Altura. These included funding from The National Heart, Lung and Blood Institute, The National Institute on Drug Abuse, The National Institute on Mental Health, and The National Institute on Alcoholism and Alcohol Abuse.
References
- Ferreira-Martins J, Leite-Moreira AF (2010) Physiologic basis and pathophysiologic implications of the diastolic properties of the cardiac muscle. BioMed Research International.
- Maeder MT, Kaye DM (2009) Heart failure with normal left ventricular ejection fraction. J Am Coll Cardiol 53(11): 905-918.
- Bursi F, Weston S, Redfield MM, Jacobsen SJ, Pakhomov S, et al. (2006) Systolic and diastolic heart failure in the community. JAMA 296(18): 2209-2216.
- Owan TE, Hodge DO, Herges RM, Jacobsen SJ, Roger VL, et al. (2006) Trends in prevalence and outcome of heart failure with preserved ejection fraction. N Engl J Med 355(3): 251-259.
- Weber M, Hamm C (2006) Role of B-type natriuretic peptide (BNP) and NT-proBNP in clinical routine. Heart 92(6): 843-849.
- Dahlstorm U (2004) Can natriuretic peptides be used for the diagnosis of diastolic heart failure? Eur J Heart Fail 6: 281-287.
- Kurl S, Ala-Kopsala M, Ruskoaho H, Makikallio T, NyyssonenK, et al. (2009) Plasma N-terminal fragments of natriuretic peptides predict the risk of stroke and atrial fibrillation in men. Heart 95(13): 1067-1071.
- Ng AC, Delgado V, Bertini M, van der Meer RW, Rijzewijk LJ, et al. (2009) Findings from Left Ventricular Strain and Strain Rate Imaging in Asymptomatic Patients With Type 2 Diabetes Mellitus. Am J Cardiol 104(10):1398-1401.
- Dokainish H, Nguyen JS, Sengupta R, Pillai M, Alam M, et al. (2010) Do additional echocardiographic variables increase the accuracy of E/e′ for predicting left ventricular filling pressure in normal ejection fraction? An echocardiographic and invasive hemodynamic study. J Am Soc Echocardiogr 23(2): 156-161.
- Calvieri C, Rubattu S, Volpe M (2012) Molecular mechanisms underlying cardiac antihypertrophic and antifibrotic effects of natriuretic peptides. J Mol Med 90(1): 5-13.
- Mishra RK, Li Y, Ricardo AC, Yang W, Keane M, et al. (2013) Association of N-Terminal pro-B-type natriuretic peptide with left ventricular structure and function in chronic kidney disease (from the Chronic Renal Insufficiency Cohort [CRIC]). Am J Cardiol 111(3): 432-438.
- Troughton R, Richards AM (2009) B-type natriuretic peptides and echocardiographic measures of cardiac structure and function. JACC Cardiovasc Imaging 2(2): 216-225.
- Yoneyama A, Koyama J, Tomita T, Kumazaki S, Tsutsui H, et al. (2008) Relationship of plasma brain-type natriuretic peptide levels to left ventricular longitudinal function in patients with congestive heart failure assessed by strain Doppler imaging. Int J Cardiol 130(1): 56-63.
- Ommen SR, Nishimura RA, Appleton CP, Miller FA, Oh JK, et al. (2000) Clinical utility of Doppler echocardiography and tissue Doppler imaging in the estimation of left ventricular filling pressures: A comparative simultaneous Doppler-catheterization study. Circulation 102(15): 1788-1794.
- Solomon SD, Verma A, Desai A, Hassanein A, Izzo J, et al. (2010) Effect of intensive versus standard blood pressure lowering on diastolic function inpatients with uncontrolled hypertension and diastolic dysfunction. Hypertension 55(2): 241-248.
- Imran U, Susan C, Chung-Leih H, Anil V, James D, et al. (2013) Relation of N-Terminal Pro-B-Type Natriuretic Peptide with Diastolic Function in hypertensive Heart Disease. Am J Hypertens 26(10): 1234-1241.
- Kasner M, Westermann D, Steendijk P, Gaub R, Wilkenshoff U, et al. (2007) Utility of Doppler echocardiography and tissue Doppler imaging in the estimation of diastolic function in heart failure with normal ejection fraction: a comparative Doppler conductance catheterization study. Circulation 116(6): 637-647.





