The Small and Powerful Nanobodies (Nbs) Originating from Peculiar Animals used for Therapy of Cancers
Zhang N1, Guo H1, Zheng W2, Wang P1, Ma X1*
1 School of Biotechnology and State Key Lab oratory of Bioreactor Engineering, East China University of Science and Technology, Shanghai, China.
2 School of Pharmacy and Shanghai Key Laboratory of New Drug Design, East China University of Science and Technology, Shanghai, China.
*Corresponding Author
Xingyuan Ma,
Professor, School of Biotechnology and State Key Laboratory of Bioreactor Engineering,
East China University of Science and Technology, Shanghai 200237,
China.
Tel/Fax: +86 2164250135
E-mail: maxy@ecust.edu.cn
Received: February 14, 2016; Accepted: March 15, 2016; Published: March 29, 2016
Citation: Zhang N, Guo H, Zheng W, Wang P, Ma X (2016) The Small and Powerful Nanobodies (Nbs) Originating from Peculiar Animals used for Therapy of Cancers. Int J Cancer Stud Res. S4:001, 1-7. doi: http://dx.doi.org/10.19070/2167-9118-SI04001
Copyright: Ma X© 2016. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
A conventional antibody is structurally featured with two heavy chains and two light ones. However, a simpler version termed as heavy-chain antibody (HCAb), also naturally exists in the serum of peculiar animal such as camels and llamas. HCAb contains a single heavy chain with one variable domain (VHH domain) and two constant domains (CH2 and CH3 domain). Interestingly, the fragment corresponding to single variable domain in HCAb can exist stably and work independently (nanobody, Nb), something like an abbreviated HCAb. Its smaller molecular weight confers Nbs unique and prominent merits including low possibility of eliciting undesirable immune response, strong tissue penetrability, high stability and ease of production. In this review, after discussing the features of Nb, we systematically reviewed the most recent advances in the wide applications of Nbs in the everlasting war against cancer. More emphasis is put on their applications as cancer therapeutics agents (ligand blocker, receptor antagonist) and as components of cancer therapeutic bioconjugates (with exotoxin, liposome, T-lymphocyte, or enzyme).
2.Abbreviations
3.Introduction
4.The Structural differences in VH and VHH
5.Characteristics of Nbs
5.1.Low Immunogenicity
5.2.Strong Tissue Penetrability
5.3.Easy Expression
6.The Strategies of Using Nb in the Fighting against Cancers
6.1.Nbs as Antagonists for Cancer Therapy
6.2.Nb as a Ligand Blocker
6.3.Nb as a Receptor Antagonist
6.4.Nbs-based Bioconjugate for Cancer Therapy
6.5.Nb-exotoxin Conjugates in Tumor Targeting
6.6.Nb-liposome Conjugates for Cancer Therapy
6.7.Nb-T lymphocyte Conjugates against the Cancer
6.8.Nb-enzyme Conjugates for Cancer Therapy
7.Conclusion
8.Acknowledgments
9.References
Abbreviations
CH: The constant domain in the heavy chain of antibodies (or of heavy-chain antibodies); HCAb: Heavychain
antibody; mAbs: Monoclonal antibodies; Nb: Nanobody; VH: The variable domain of the heavy chain of antibodies; VHH: The variable domain of the heavy chain of heavy-chain antibodies; VL: The variable domain of the light chain of antibodies.
Keywords
Single-Domain Antibody; Immunogenicity; Penetrability and Stability; Therapeutic Anticancer Bioconjugates.
Introduction
Köhler and Milstein reported the hybridoma technology that could be used to efficiently produce monoclonal by mouse in 1975 [1]. Similar to the concept of magic bullets imagined by Paul Ehrlich at the beginning of the 20th century [2], it has been widely believed that such a molecule would be ideal for detection therapy. Indeed, mAbs-based therapy is one of the most and important strategies for treating patients with haematological malignancies or solid tumors. Currently, there are six mAbs that have been approved for applications in cancer by the Food and Drug Administration (FDA) of USA [3]. MAbs have been selected for their high specificity and affinity to the protein biomarkers related to cancer. Thanks to the acceptable stability of mAbs, they are hailed as the right agent to be used as target molecules. However, such a “nice reagent” is inevitably facing serious problems. The most pronounced one should be the undesired immune response due to the intrinsic immunogenicity resulted from their murine origin, frequently leading to the rejection of immune system by the administrated patients. Although humanization provides a hopeful possible solution to the side-effect, it is also a time-and energy-consuming huge task, because the conventional mAbs is really large. For the same reason, the poor pharmacokinetics and weak tissue penetration makes more than 80% of the administered mAbs typically useless regarding the interaction with tumors [4]. The low efficacy of mAbs in clinical practice renders the production expensive, implying the deathly need for further improvement of mAbs in cancer treatment.
It is as urgent as the finding of therapy against cancer to discover new antibodies that provide the same binding specificity and affinity comparable to mAbs. Recently, some engineered smaller recombinant fragments (e.g. antigen-binding fragment, Fab, ~50 kDa; variable fragment, Fv, ~15 kDa; single-chain variable fragment, ScFv, ~30 kDa), were extensively tested in the hope of overcoming the drawbacks of the full-length mAbs [5, 6]. Nevertheless, their wide applications are severely limited due to lower affinity, weak stability, production difficulty, vulnerability to proteinase and the tendency to aggregation.
By serendipity, a special type of naturally existing antibody was discovered in animals from the Camelidae family by Hamers- Casterman and coworkers in 1993 [7]. It is essentially a kind of heavy-chain antibody (HCAb), which consists of one VHH (variable domain, responsible for antigen binding), and two constant domains (CH2 and CH3). Despite the absence of light chain and the first constant domain (CH1), HCAbs can bind to their antigens with similar affinity to that of conventional antibodies [8]. More importantly, when the part corresponding to VHH is independently cloned and expressed, a polypeptide capable of binding antigen will be obtained (Figure 1). It provides a great convenience in desirable antibody production. Since the size of VHH antibody is at the scale of nanometer. (Diameter of 2.5 nm and height of 4 nm), VHH was renamed as nanobody (Nb) [9, 10], or alternatively aliased single domain antibody (sdAb) [11]. Due to the wide adoption of the term nanobody, we use it though out the text. So far, in addition to the DNA or RNA aptamers, Nb is the smallest known functional antigen-binding fragment as Table 1. In a word, in terms of functionality, Nb is more than a conventional antibody.
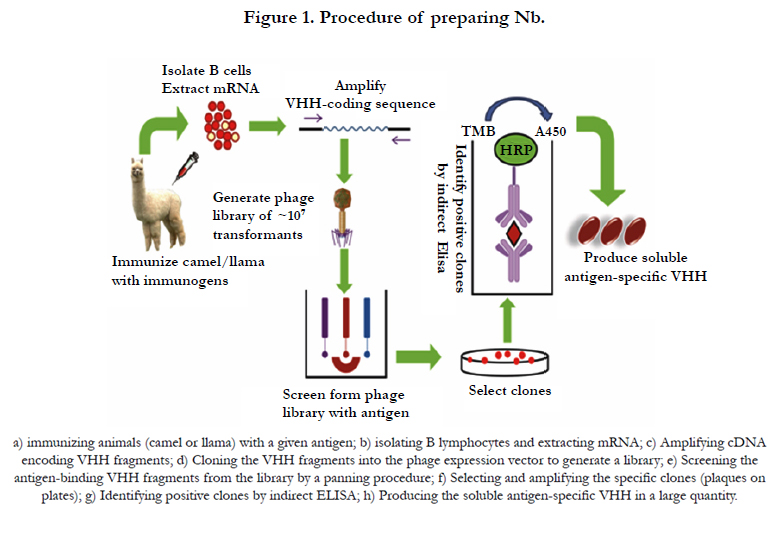
Figure 1. Procedure of preparing Nb.
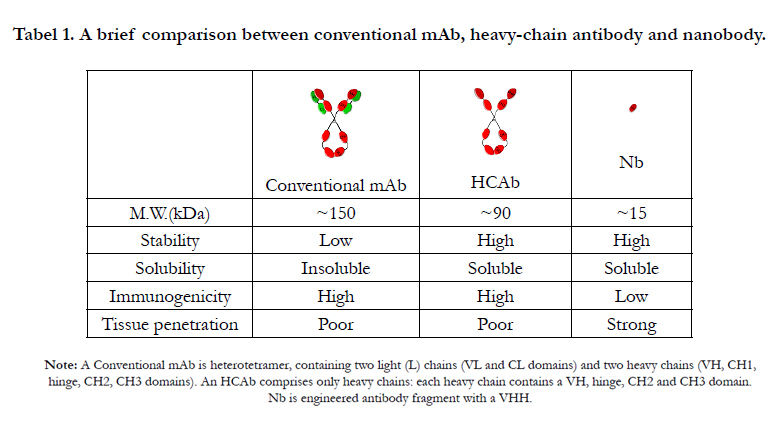

Tabel 1. A brief comparison between conventional mAb, heavy-chain antibody and nanobody.
Note: A Conventional mAb is heterotetramer, containing two light (L) chains (VL and CL domains) and two heavy chains (VH, CH1, hinge, CH2, CH3 domains). An HCAb comprises only heavy chains: each heavy chain contains a VH, hinge, CH2 and CH3 domain. Nb is engineered antibody fragment with a VHH.
The Structural differences in VH and VHH
The crystal and solution structures [12-16] of several nanobodies have been solved and saved in database. It is shown that their scaffolds consist of two α-sheet structures similar to conventional mAbs [17]. The remarkably similar organization lies in the variable domain of the H-chain (VH) of a mAb and that of an HCAb (VHH) [18]. Nevertheless the VH and VHH contain subtle but important differences that can explain the antigenbinding capacity of the VHH in a single-domain format. Main differences are in framework-2 (FR2) and in complementarity determining regions (CDRs). Specifically, within FR2, four highly conserved hydrophobic amino acids that normally participate in the interaction with the VL domain [19] are replaced in a VHH by four hydrophilic amino acids (Phe37, Glu44, Arg45, and Gly47) [20-22] (Figure 2). Some experiments proved that a mouse VH in absence of the VL partner is sticky (tends to aggregates), but the substitutions of FR2 to mimic the amino acids of camelid VHH renders the camelized domain more soluble [23]. Hence, the substitutions of these amino acids are frequently used to explain the increased solubility of Nbs. In VHH, the antigen is recognized by only three loops instead of six loops; however, the loops are longer in a VHH than in a VH of a conventional mAb. In VHH, the antigen is recognized by only three loops instead of six loops; however, the loops are longer in a VHH than in a VH of a conventional mAb. It has been proposed that the VHH paratope is enlarged by an extension of the CDR1, and especially the CDR3 [24]. The CDR1 that normally involves residues 31-35 in all VH is entended to encompass residues 27-35 in camelid VHH. The VHH-specific hypervariable residues 27-30, forming the loop connecting the two β-sheets of the immunoglobulin fold, are exposed to solvent. The average length of CDR3 observed in human VH is 13 amino acids and around 18 in VHH [25, 26]. A longer CDR3 in Nb allows it to form a finger-like structure capable of extending into cavities on target proteins, enabling Nb to bind to unique epitope [27, 28]. The larger CDRs of the VHH will widen the actual surface of the antigen binding site, which might compensate for the absence of the antigen-binding surface area provided by the VL domain. By contrast, the binding interfaces of Fab-and other mAbs-derived fragments are flatter and less flexible, rendering the interactions of mAbs and fragment derivatives solely confining to the surface of antigens [29]. The prolate (rugby ball–shaped) structure of the VHH domain forms a convex paratope surface, which makes it extremely suitable to insert into a cavity on the surface of the antigen [30]. The small size of the Nb allows the epitope recognition even when the target proteins are densely packed [31]. Compared with the VH, many VHH sequences contain a known interloop disulfide bond from an extra pair of cysteine residues in the CDR1 and CDR3, helping to improve stability by restricting the flexibility of the loops in Nb when an antigen is absent.

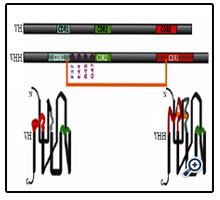
Figure 2. The sequence organization of the VH and VHH [32].
Characteristics of Nbs
Common mAbs have many vital features, such as high affinity and specificity to a target molecule (protein or other molecule), high sensitivity and low inherent toxicity. In addition to these merits, Nbs uniquely possesses other technological and biophysical advantages that enable them to outperform conventional counterpart in several facets.
The majority of mAbs are produced with a murine origination, and they are not perfectly compatible with the human immune system, thus restricts their biological efficacy. By contrast, Nbs tested to date do not show any intolerable level of immunogenicity in preclinical and clinical settings. This is not surprising when the facts that about 90% frameworks are identical with human VH domains, and that additional engineering is usually performed to further humanize the Nbs for use in the clinic, are taken into account. Now, a strategy has been developed to humanize the 12 out of a total of 14 amino acids that differ in human VH and camelid VHH [33].
Poor tissue penetrability inherent to mAbs is a serious obstacle to the further development of antibody therapeutics, in particular, for immunotherapy of solid tumors [34]. Although 85% of human cancers involve solid tumors, only 3 out of 10 mAbs approved in the US for cancer therapy are directly targeting antigens highly expressed on the surface of solid tumors (Vectibix and Erbitux targeting EGF and Herceptin targeting Her2) [35]. This disproportion reflects the difficulty in reaching an effective concentration of antibodies in solid tumors due to the poor tissue penetrability of conventional mass. Nbs have been great potential to overcome this problem because of the smaller size. In addition, rapid clearance from the blood also requires great penetrating power as a plausible feature for desired clinic utilization.
While the production of recombinant conventional mAbs is often hampered by aggregation and reduced affinity from mispairing of the VH and VL domains in mammals, production of multivalent Nbs is straightforward; the lack of necessary post-translational modifications allows the production of Nbs in bacteria or yeast [36]. Nbs can be expressed with microbial expression systems such as Escherichia coli, Saccharomyces cerevisiae, Pichia pastoris and filamentous fungi. Microbial expression promise an obvious advantage and possibility for the extensive use of Nbs because the average cost is 2–3 fold less compared with mammalian expression system. A high yield of 250 mg/L in expression of the binding domain (VHH) in S. cerevisiae at shaking flask scale has been reported [37].
The Strategies of Using Nb in the Fighting against Cancers
Targeted therapy of cancer requires the drug to reach the site of disease in the body rapidly, and promise a minimum of damage to normal tissue. At present, a variety of antibody drugs have been studied for the treatment of cancer. However, these antibodies are large protein, making it is difficult to penetrate into tumor cells. Their complicated structures also make it rather difficult and expensive to produce. However, Nbs are advantageous in this aspect. Therefore, it is not surprising to see that Nbs are more suitable for treatment in tumors than other antibodies as Table 2.
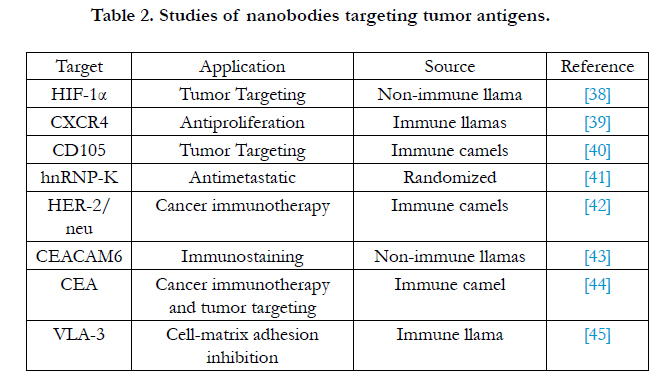
Table 2. Studies of nanobodies targeting tumor antigens.
A significant feature of Nb technology is the versatility of the system in enabling the production of a wide variety of Nbs with specificity to different targets. For example: there are two possible strategies to inhibit growth of tumor: using Nbs as ligand blockers or antagonists of the receptor. The anti-ligand Nbs can be efficiently used only when a single ligand is involved in the signaling of the corresponding receptor. For instance, while EGFR can be activated by different ligands, the hepatocyte growth factor (HGF) is the only known ligand for the HGF receptor, i.e. c-Met [46] (Figure 3A).
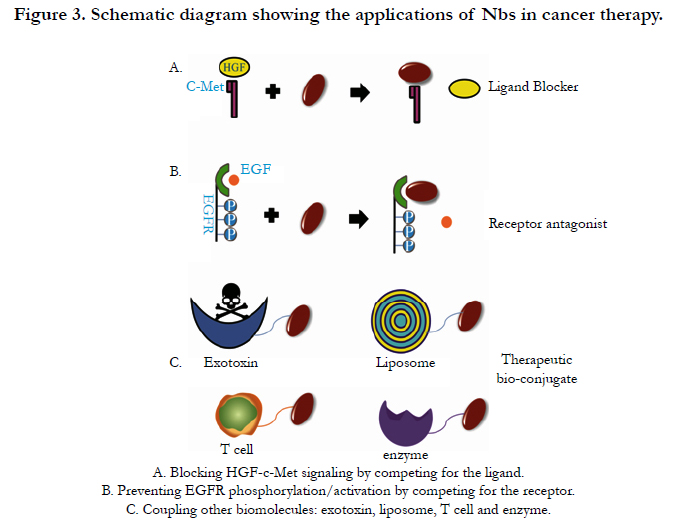
Figure 3. Schematic diagram showing the applications of Nbs in cancer therapy.
Hepatocyte growth factor (HGF) is a plasminogen-like protein that is secreted by mesenchymal cells as single inactive peptides and c-Met is the tyrosine kinase receptor of HGF. HGF-c-Met signaling is involved in many human malignancies. In tumor cells where HGF and c-Met are overexpressed, the enhanced signaling can trigger tumorogenesis and angiogenesis, and thus can prevent apoptosis, which all contribute to the outgrowth of tumors [47]. Onartuzumab, a monovalent antibody against c-Met, which has been shown to significantly improve both progressionfree survival and overall survival in c-Met-positive patients with NSCLC when used in combination with the epidermal growth factor receptor (EGFR) inhibitor erlotinib [48]. Notably, Tobias et al. examined the anticancer effects of the anti-c-Met Nb (from Ablynx NV) in vitro using human multiple myeloma (MM) as a model system. This novel drug showed high specificity and potency for blocking HGF-c-Met signaling in human myeloma cell lines, and completely blocked HGF induced thymidine incorporation, migration, and adhesion in multiple myeloma cell lines, reversing the HGF-induced inhibition of osteoblastogenesis. They also compared the Nb with anti-c-Met mAbs and revealed the similar or better effect. So the applicability of anti-c-Met Nb can be a valuable novel treatment option for MM and other cancers driven by HGF-c-Met signaling [49].
Epidermal growth factor receptor (EGFR) is deemed to be one of the main molecular targets for treatment of cancer. It has been identified that EGFR involves in pathogenesis of some forms of human cancers. Although Anti-EGFR mAbs have shown very impressive results at the preclinical level, results have not always live up to the expectations in the clinic, which is only partly due to the difficulties in assessing EGFR expressions status in patients [50]. Oliveira et al. described the use of functional screening of antibody phage library using competitive elution with the ligand EGF to develop antagonistic anti-EGFR Nbs for cancer therapy (Figure 3B). In an in vivo murine xenograft model, the Nbs were effective in delaying the outgrowth of A431-derived solid tumours. The identified antibody fragments were found to efficiently inhibit binding of EGF to the EGFR without acting as receptor agonists themselves. In addition, they blocked EGFmediated signaling and EGF-induced cell proliferation [51].
The strict monomer structure and small size of Nbs provide the significant convenience in making multiple functions conjugates with other partners. The design of fusion constructs of Nbs and other functional modules provides a strategy for the development of novel cancer therapeutics (Figure 3C).
Angiogenesis plays a crucial role in the growth, invasion and metastasis of cancer. Vascular endothelial growth factor receptor 2 (VEGFR2) is such a significant target that is overexpressed in tumor vasculature compared with the endothelium cells of resting blood vessels. Blocking of its signaling can inhibit neovascularization and tumor metastasis. Immunotoxins represent a promising group of targeted therapeutics to combat tumors [52]. Behdani et al. fused a VEGFR2-specific Nb to the truncated form of Pseudomonas exotoxin A and evaluated its ability to bind the VEGFR2 on the cell surface. They demonstrated that the immunotoxin inhibited the proliferation of VEGFR2-expressing cells in vitro [53]. This finding provided a novel strategy for tumor therapy.
Different targeting moieties have been grafted to liposomes’ surface [54]; therefore, liposomes can be successfully employed as a carrier system for a variety of molecules. Oliveira et al. tested a new multivalent platform, which consisted of polyethylene glycol (PEG) –liposomes coupled to Nb that recognized the ectodomain of EGFR (EGa1). They investigated EGFR internalization and down-regulation of this system in vitro and in vivo, and they found that EGa1-L induced a significant inhibition of tumor cell proliferation in vitro due to the effect likely mediated by the combination of receptor down-regulation and receptor antagonism. Also an in vivo down-regulation of EGFR was observed in tumors of mice intravenously injected with EGa1-L, indicating that this multivalent platform blocks ligand binding to the receptor and simultaneously initiated the regulation [53].
T lymphocyte (or T cell) plays many key roles in controlling and combating cancers. Tumor cells deploy a variety of immuneevasion strategies to escape elimination by T cells. To circumvent this problem, genetic engineering of T cells was used to generate the supernatural T cells (T-body) which express chimeric antigen receptor (CAR) and possess attractive capabilities. CARs are typically composed of an extracellular antibody recognition domain specific to tumor antigen, a transmembrane and intracellular signaling domains linked by a hinge region [56-58]. Nb with desirable characteristics was utilized to generate engineered T cell [59]. Rahbarizadeh et al. used the anti-MUC1 (a type I transmembrane protein, significantly overexpressed and aberrantly glycosylated in tumors of epithelial origin) Nb gene as an antigen binding fragment in an Nb-T lymphocyte conjugate for cancer therapy [60]. The results showed that this T-body specifically targeted tumor cells, and the strategy has broad prospects for cancer therapy.
Carcinoembryonic antigen (CEA) is highly expressed on cancer cells of epithelial origin, such as colorectal, lung, breast, and ovarian carcinoma [61]. It is not expressed in other cells of the body except for low-level expression in gastrointestinal tissue, which makes it an attractive target for tumor therapy. Recently, Cortez- Retamozo and co-workers identified an Nb with sub-nanomolar affinity to the human tumor-associated carcinoembryonic antigen. When the Nb was conjugated to lactamase from Enterobacter cloacae, and its capacity in site-selectively activating anticancer prodrug was evaluated. The conjugate was readily purified in high yields without aggregation or loss of functionality of each constituent. In vitro experiments showed that, the Nb–enzyme conjugate can effectively activate the release of phenylenediamine mustard from the cephalosporin nitrogen mustard prodrug (7-(4-carboxybutanamido) cephalosporin mustard), at the surface of carcinoembryonic antigen-expressing LS174T cancer cells. In vivo studies also demonstrated that the conjugate had an excellent biodistribution profile and induced regressions and cures of established tumor xenografts. The easy generation and manufacturing at reasonable yield of Nb-based conjugates together with their potent antitumor activity make Nb a promising vehicle for a new generation of cancer therapeutics [62].
Conclusion
Nbs have some unique desirable properties such as smaller size, superior solubility, intrinsic stability, relatively easy engineering into multiple potent constructs, highly capable in recognizing uncommon or even hidden epitopes (and binding into cavities or active sites of enzyme targets), ease of manufacture. All these characteristics imply the remarkable promise that, in paralleling with conventional antibodies, Nbs can be exploited as robust cancer therapeutic reagents with novel or fully tested strategies, providing new hope of the final win in the combating against health-threating cancers.
Acknowledgments
This study was supported by the National Natural Science Foundation (30873190, 31300660), Science and technology innovation action plan of Shanghai (14431904300), Shanghai Pujiang Program (13PJD012), and a foundation for young teacher from Education Ministry of China (20120074120027)and partially supported by Open Funding Project of the State Key Laboratory of Bioreactor Engineering.
References
- Köhler, G., Milstein, C (2005) Continuous cultures of fused cells secreting antibody of predefined specificity. 1975. J Immunol 256(5517): 495-497.
- ElBakri A, Nelson PN, Abu Odeh RO (2010) The state of antibody therapy. Hum Immunol 71(12): 1243-1250.
- Chames P, Van Regenmortel M, Weiss E, Baty D (2009) Therapeutic antibodies: successes, limitations and hopes for the future. Brit J Pharmacol 157(2):220-233.
- Beckman RA, Weiner LM, Davis HM (2007) Antibody constructs in cancer therapy: protein engineering strategies to improve exposure in solid tumors.Cancer 109(2): 170-179.
- Gong R, Chen W, Dimitrov DS (2012) Expression, purification, and characterization of engineered antibody CH2 and VH domains. Methods Mol Biol 899: 85-102.
- Miller KR, Koide A, Leung B, Fitzsimmons J, Yoder B, et al.(2012) T cellreceptor-like recognition of tumor in vivo by synthetic antibody fragment. PloS one7(8): e43746.
- Hamers-Casterman C, Atarhouch T, Muyldermans S, Robinson G, Hammers C, et al.(1993) Naturally occurring antibodies devoid of light chains. Nature 363(6428): 446-448.
- Conrath KE, Wernery U, Muyldermans S, Nguyen VK (2003) Emergence and evolution of functional heavy-chain antibodies in Camelidae. Dev Comp Immunol 27(2): 87–103.
- Tillib SV (2011) “Camel nanoantibody” is an efficient tool for research, diagnostics and therapy. Mol Biol 45(1): 66-73
- Muyldermans S (2013) Nanobodies: natural single-domain antibodies. Annu Rev Biochem 82: 775-797.
- Arbabi Ghahroudi M, Desmyter A, Wyns L, Hamers R, Muyldermans S (1997) Selection and identification of single domain antibody fragments from camel heavy-chain antibodies. FEBS Lett 414(3): 521-526.
- Vincke C, Loris R, Saerens D, Martinez-Rodriguez S, Muyldermans S, et al (2009) General Strategy to Humanize a Camelid Single-domain Antibody and Identification of a Universal Humanized Nanobody Scaffold. J Biol Chem 284(5): 3273-3284.
- Spinelli S, Frenken L, Bourgeois D, de Ron L, Bos W, et al. (1996) The crystal structure of a llama heavy chain variable domain. Nat Struct Mol Biol 3(9): 752-757.
- Decanniere K, Desmyter A., Lauwereys M, Gharoudi, MA, Muyldermans S (1999) A single-domain antibody fragment in complex with RNase A: non-canonical loop structures and nanomolar affinity using two CDR loops. Structure 7(4): 361-370.
- Renisio JG, Pérez J, Czisch M, Guenneugues M , Bornet O, et al. (2002) Solution structure and backbone dynamics of an antigen‐free heavy chain variable domain (VHH) from Llama. Proteins 47(4): 546-555.
- Vranken W, Tolkatchev D, Xu P , Tanha J, Chen Z, et al. (2002) Solution structure of a llama single-domain antibody with hydrophobic residues typical of the VH/VL interface. Biochemistry 41(27): 8570-8579.
- Deffar K., Shi H , Li L, Wang X, Zhu X (2009) Nanobodies-the new concept in antibody engineering. Afr J Biotechnol 8(12):2645-2652.
- Nguyen VK, Hamers R, Wyns L, Muyldermans S (2000) Camel heavy‐chain antibodies: diverse germline VHH and specific mechanisms enlarge the antigen‐ binding repertoire.EMBO J 19(5), 921-930.
- Chothia C, Novotny J, Bruccoleri R, Karplus M (1985) Domain association in immunoglobulin molecules: the packing of variable domains. J Mol Biol 186(3): 651-663.
- Muyldermans S, Atarhouch T, Saldanha J, Barbosa J, Hamers R (1994) Sequence and structure of V-Hdomain from naturally-occurring camel heavychain immunoglobulins lacking light-chains. Protein Eng 7(9): 1129-1135.
- Pereira SS, Moreira-Dill LS, Morais MS, Prado ND, Barros ML, et al. (2014) Novel Camelid Antibody Fragments Targeting Recombinant Nucleoprotein of Araucaria hantavirus: A Prototype for an Early Diagnosis of Hantavirus Pulmonary Syndrome, PloS one, 9(9): e108067.
- Harmsen MM, Ruuls RC, Nijman IJ, Niewold TA, Frenken LGJ, et al. (2000) Llama heavy-chain V regions consist of at least four distinct subfamilies revealing novel sequence features. Mol. Immunol 37(10): 579-590.
- Davies J, Riechmann L(1994) ‘Camelising’human antibody fragments: NMR studies on VH domain. FEBS Lett 339(3): 285-290.
- Muyldermans S (2001) Single domain camel antibodies: current status. Rev Mol Biotechnol 74(4): 277-302.
- VU KB, Gharhoudi M, Wyns L, Muyldermans S(1997) Comparison of llama VH sequences from conventional and heavy chain antibodies. Mol. Immunol 34(16): 1121-1131.
- Muyldermans S, Cambillau C, Wyns L (2001) Recognition of antigens by single-domain antibody fragments: the superfluous luxury of paired domains.Trend Biochem. Sci 26(4): 230-235.
- Wesolowski J, Alzogaray V, Reyelt J, Unger M, Juarez K, et al. (2009) Single domain antibodies: promising experimental and therapeutic tools in infection and immunity. Med. Microbiol. Immunol 198(3): 157-174.
- Schmitz KR, Bagchi A, Roovers RC, van Bergen en Henegouwen PM, Ferguson KM (2013) Structural Evaluation of EGFR Inhibition Mechanisms for Nanobodies/VHH Domains. Structure 21(7): 1214-1224.
- Desmyter A, Transue TR, Ghahroudi MA, Thi MHD, Poortmans F, et al. (1996) Crystal structure of a camel single-domain VH antibody fragment in complex with lysozyme. Nat Struct Mol Biol 3(9): 803-811.
- De Genst E, Silence K, Decanniere K, Conrath K, Loris R, et al. (2006) Molecular basis for the preferential cleft recognition by dromedary heavy-chain antibodies. P Natl Acad Sci 103(12): 4586-4591.
- Revets H, De Baetselier P, Muyldermans S (2005) Nanobodies as novel agents for cancer therapy. Expert Opin Biol Th 5(1): 111-124.
- Muyldermans S, Baral TN, Retamozzo VC, De Baetselier P, De Genst E, et al., (2009) Camelid immunoglobulins and nanobody technology. Vet. Immunol.Immunopathol 128(1): 178-183.
- Vincke C, Loris R, Saerens D, Martinez-Rodriguez S, Muyldermans S, et al. (2009) General strategy to humanize a camelid single-domain antibody and identification of a universal humanized nanobody scaffold. J Biol Chem 284(5): 3273-3284.
- Ackerman ME, Pawlowski D, Wittrup KD (2008) Effect of antigen turnover rate and expression level on antibody penetration into tumor spheroids. Mol Cancer Ther 7(7): 2233-2240.
- Kolkman JA, Law DA (2010) Nanobodies–from llamas to therapeutic proteins. Drug. Discov. Today7(2): e139-e146.
- Harmsen MM, De Haard HJ (2007) Properties, production, and applications of camelid single-domain antibody fragments. Appl Microbiol Biot 77(1): 13-22.
- Frenken LG, van der Linden RH, Hermans PW, Bos JW, Ruuls RC, et al. (2000) Isolation of antigen specific llama VHH antibody fragments and their high level secretion by Saccharomyces cerevisiae. J Biotechnol 78(1):11-21.
- Groot AJ, Verheesen P, Westerlaken EJ, Gort EH, van der Groep P, et al. (2006) Identification by phage display of single-domain antibody fragments specific for the ODD domain in hypoxia-inducible factor 1alpha. Lab Invest 86(4): 345-356.
- Jähnichen S, Blanchetot C, Maussang D, Gonzalez-Pajuelo M, Chow KY, et al., (2010) CXCR4 nanobodies (VHH-based single variable domains) potently inhibit chemotaxis and HIV-1 replication and mobilize stem cells. P Natl Acad Sci 107(47): 20565-20570.
- Ahmadvand D, Rasaee MJ, Rahbarizadeh F, Kontermann RE, Sheikholislami F. (2009) Cell selection and characterization of a novel human endothelial cell specific nanobody. Mol. Immunol 46(8): 1814-1823.
- Inoue A, Sawata SY, Taira K, Wadhwa R. (2007) Loss-of-function screening by randomized intracellular antibodies: Identification of hnRNP-K as a potential target for metastasis. P. Natl. Acad. Sci. 104(21): 8983-8988.
- Sheikholeslami F, Rasaee MJ, Shokrgozar MA, Dizaji MM, Rahbarizadeh F, et al. (2010) Isolation of a novel nanobody against HER-2/neu using phage displays technology. Lab Med. 41(2): 69-76.
- Mai KT, Perkins DG, Zhang J, Mackenzie CR. (2006) ES1, a new lung carcinoma antibody—an immunohistochemical study. Histopathology 49(5): 515-522.
- Cortez-Retamozo V, Backmann N, Senter PD, Wernery U, De Baetselier P, et al. (2004) Efficient cancer therapy with a nanobody-based conjugate. Cancer Res 64(8): 2853-2857.
- Groot AJ, Khattabi ME, Sachs N, van der Groep P, van der Wall E, et al. (2009) Reverse proteomic antibody screening identifies anti adhesive VHH targeting VLA-3. Mol. Immunol. 46(10): 2022-2028.
- Reznik TE, Sang Y, Ma Y, Abounader R, Rosen E, et al. (2008) Transcription- dependent epidermal growth factor receptor activation by hepatocyte growth factor. Mol. Cancer Res. 6(1): 139-150.
- Boccaccio C, Comoglio PM. (2006) Invasive growth: a MET-driven genetic programme for cancer and stem cells. Nat. Rev. Cancer 6(8): 637-645.
- Spigel DR, Ervin TJ, Ramlau R, Daniel DB, Goldschmidt JH, et al. (2011) Final efficacy results from OAM4558g, a randomized phase II study evaluating MetMAb or placebo in combination with erlotinib in advanced NSCLC. J. Clin. Oncol. 29(suppl 15): 7505.
- Schmidt Slørdahl T, Denayer T, Helen Moen S, Standal T, Børset M, et al. (2013) Anti-c-MET Nanobody®–a new potential drug in multiple myeloma treatment. Eur. J. Haematol. 91(5): 399-410.
- Pantaleo MA, Nannini M, Fanti S, Boschi S, Lollini PL, et al. (2009) Molecular imaging of EGFR: it's time to go beyond receptor expression. J. Nucl. Med. 50(7): 1195-1196.
- Roovers RC, Laeremans T, Huang L, De Taeye S, Verkleij AJ, et al. (2007) Efficient inhibition of EGFR signalling and of tumour growth by antagonistic anti-EGFR Nanobodies. Cancer Immunol. Immun. 56(3): 303-317.
- Folkman J. (2007) Angiogenesis: an organizing principle for drug discovery?. Nat. Rev. Drug. Discov. 6(4): 273-286.
- Behdani M, Zeinali S, Karimipour M, Khanahmad H, Schoonooghe S, et al. (2013) Development of VEGFR2-specific Nanobody Pseudomonas exotoxin A conjugated to provide efficient inhibition of tumor cell growth. New Biotechnol. 30(2): 205-209.
- Torchilin VP. (2005) Recent advances with liposomes as pharmaceutical carriers. Nat. Rev Drug Discov 4(2): 145-160.
- Oliveira S, Schiffelers RM, van der Veeken J, van der Meel R, Vongpromek R, et al. (2010) Downregulation of EGFR by a novel multivalent nanobodyliposome platform. J. Control. Release 145(2): 165-175.
- Friedmann-Morvinski D, Bendavid A, Waks T, Schindler D, Eshhar Z. (2005) Redirected primary T cells harboring a chimeric receptor require costimulation for their antigen-specific activation. Blood 105(8): 3087-3093.
- Sadelain M, Brentjens R, Rivière I. (2009) The promise and potential pitfalls of chimeric antigen receptors. Curr. Opin. Immunol. 21(2): 215-223.
- Sadelain M, Rivière I, Brentjens R. (2003) Targeting tumours with genetically enhanced T lymphocytes. Nat. Rev. Cancer 3(1): 35-45.
- Bakhtiari SHA, Rahbarizadeh F, Hasannia S, Ahmadvand D, Iri-Sofla FJ,et al. (2009) Anti-MUC1 nanobody can redirect T-body cytotoxic effector function. Hybridoma 28(2): 85-92.
- Rahbarizadeh F, Ahmadvand D, Sharifzadeh Z. (2011) Nanobody; an old concept and new vehicle for immunotargeting. Immunol. Invest. 40(3):299-338.
- Hammarström S. (1999) The carcinoembryonic antigen (CEA) family: structures, suggested functions and expression in normal and malignant tissues. Semin. Cancer Biol. 9(2): 67-81.
- Cortez-Retamozo V, Backmann N, Senter PD, Wernery U, De Baetselier P, et al. (2004) Efficient cancer therapy with a nanobody-based conjugate.Cancer Res. 64(8): 2853-2857.





