28-Homocastasterone: A Novel Plant Ketosteroid Enhances Tissue Transaminase Catalytic Activity and Gene Expression in Normal and Diabetic Male albino wistar Rat
Athithan V, Srikumar K*
Department of Biochemistry and Molecular Biology, School of Life Science, Pondicherry University, Pondicherry, India.
*Corresponding Author
Kotteazeth Srikumar M.Sc., Ph.D., FABMS,
Professor, Department of Biochemistry and Molecular Biology,
School of Life Science, Pondicherry University, Pondicherry, India.
Tel: 0413-2654422
Fax: 0413-2654422
E-mail: ketoathi@gmail.com
Received: June 26, 2016; Accepted: July 15, 2016; Published: July 20, 2016
Citation: Athithan V, Srikumar K (2016) 28-Homocastasterone: A Novel Plant Ketosteroid Enhances Tissue Transaminase Catalytic Activity and Gene Expression in Normal and Diabetic Male albino wistar Rat. Int J Clin Pharmacol Toxicol. 5(5), 220-224.DOI : dx.doi.org/10.19070/2167-910X-1600037
Copyright: Srikumar K© 2016. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
28-Homocastasterone (28-HC) is a brassinosteroid family phytohormone. Humans were exposed through diet and herbal based folk medicine, although its mammalian cellular influence unknown. Present study investigated 28-HC cellular metabolic enzyme inductivity on tissues in normal and diabetic rats. Induction of diabetes was achieved by single peritoneal injection of STZ (60mg/kg.bwt) followed by 28-HC were feed orally 15 days, daily. At end of 15th day tissues was removed followed by enzyme activity and mRNA expression of alanine transaminase (ALT), aspartate transaminase (AST), alkaline phosphatase (ALP) was analysed. 28-HC treated rat tissues ALT, AST, GGT and ALP catalytic activity and mRNA expression increased significantly (P<0.05). Dietary plant ketosteroid 28-HC augmented transaminase and phosphatase activity in tissues resulted to cellular transamination and phosphorylation status affected by this plant steroid in rat. It is suggested that 28-HC supplemented diets may be suitable for higher energy related work activities in man and animals.
2.Introduction
3.Materials and Method
3.1.Materials
3.2.Experimental animal
3.3.Induction of diabetes and 28-HC administration
3.4.Tissue homogenate preparation
3.5.Analysis of enzymes activity
3.6.RNA isolation and mRNA expression analysis
3.7.Statistical analysis
4.Results
5.Discussion
6.Conclusion
7.Acknowledgement
8.References
Keywords
28-Homocastasterone; Phytoketosteroid; Transaminase; Phosphatase; Rat Tissues.
Introduction
Cellular metabolic processes are driven by metabolite availability, thermodynamic considerations and catalytic activities. Homeostatic regulatory processes are built into cell and tissue function to ensure equilibrium dynamics in cell function and includes factors such as rate limiting steps, circulatory partitioning, active/allosteric /product inhibition, coenzyme role, protein synthesis and gene expression events. Modulatory influences are recognized by living cells through physico-chemical means and cell signaling cascades. Environmental and epigenetic contributions are therefore known to trigger metabolic changes eventually affecting the genetic apparatus.
Diet being a major resourceful environmental factor dietary constituents exhibit a significant role for modulating cellular metabolic events on time scales of seconds to hours. Since dietary factors in general are natural products, it has become essential to investigate the effect/role of known dietary natural compounds for their cellular metabolic influences, thus acquising early translational knowledge on the merits and demerits of use of such compounds as potential therapeutic interventions for the future. Since globally, the science of natural products has attracted greater attention as an alternative to the toxic end results noted following long-term use of allopathic therapeutics, and since India had an established legacy for use of herbal drugs for managing health conditions, dating back to 2500 BC under the umbrella of Ayurveda, modernizing the ritualistic practice of Ayurveda through analytical logic supported with modern scientific and instrumental approaches would creditably revalidate and re-establish the credibility of this system of Indian medical practice, the world over [1].
In order to make a meaningful beginning in this direction, studies employing plant steroids were carried out in this laboratory, and as a part of which this work was independently initiated to understand the biological effectiveness of use the phytosteroid 28-homocastasterone (28-HC) in mammalian cell biology employing male wistar rat as a model [2, 3].
Liver known to be central organ for carbohydrate metabolism, lipid and protein synthesis, liver enzymes ALT and AST have link carbohydrates, lipid and protein metabolism. Increase in ALT enzyme activity suggests increase transamination of the amino acid alanine to α-ketoglutarate, a reactive intermediate in the crabs TCA cycle leading to the formation of pyruvate [4]. Elevated pyruvate levels in a cell contribute to bio energy production through the formation of ATP in the cells of tissues. However reduced tissues ALT activity suggest that bio energetics exist negative to that in tissues [5]. Assessment of the AST enzyme activity indicated transamination between aspartate and glutamate contributing to the reduced generation of alpha-ketoglutarate and oxaloacetate [6]. Perturbations affecting these enzymes catalytic activates and transcription in the animal tissues signaling alteration metabolic process involving them. Such perturbations were therefore indicator of cell stress. 28-HC exhibited structural similarity with animal steroid hormones, they had cellular nuclear receptors to communicate their biological responses. Dietary phyto ketosteroid 28-HC can therefore suspected of metabolic stress in animal tissues [7]. Investigation were therefore directed to elucidated the bio-molecular effect of the 28-HC, on the ALT, AST, GGT and ALP catalytic and transactivation in normal and diabetic male albino wistar rat.
Materials and Method
Chemicals used in this study were purchased from sigma Aldrich (USA) were analytical grade. The reagents and enzymes kit were purchased from agape diagnostic, Kerala, India. Test compound 28-Homocastasterone was gifted by Dr.V.S.Pori, National Chemical Laboratory (CSIR), Pune, India. Double distilled water was used to prepare all the regents in this study.
The laboratory animal used in this study, albino wistar rats, male and two week old (150-180 body weight range), were purchased from Sri Ragavendra Enterprises, Bangalore, India. Experiment protocol approval obtained from institutional animal ethics committee, Pondicherry University, India (Aproval.No.IAEC/2013/01). The animals handling protocol strictly followed as per the guidelines of the CPCSEA. Rats were housed in hygienic and free air circulation condition and allowed to freely access normal rat chows and water ad libitum throughout experimental period, for acclimation animal room temperature were maintained at 25°C and given 12 hours light and dark cycle. After one week acclimation, 24 healthy and average weight 150gm body weight, three week old male rats were selected in this study. Rats was randomly divided into four groups each group contain six rats (n=6) such as,
Group I: Normal control (50% ethanol 0.1mL)
Group II: Normal +100μg 28-HC in 50% ethanol 0.1 mL
Group III: Diabetic control (50% ethanol 0.1mL)
Group IV: Diabetic + 100μg 28-HC in 50% ethanol 0.1 mL
The single peritoneal injection of STZ (60 mg/kg.bwt in 0.1M citrate buffer pH 4.5) was made at overnight fasted rats. After 48 hours blood glucose level was cheeked with help of glucometer (accuva cheek) in blood. The blood glucose >250 mg/ dl were considered to be a diabetic used in this study. Group II and IV were treated with 28-homocastasterone 666.66μg/kg.bwt (100μgm) for 15 consecutive days, daily and group I and III serve as normal and diabetic control received 50% ethanol alone [2].
At the end of 15th day of treatment rats were killed and tissues liver, kidney and testis were immediately removed and washed with phosphate buffer (0.1M, pH 7.4), stored at -80°C until used for assay. 1 gm of tissue weight and homogenized in phosphate buffer with help of motorized tissue homogenizer (Teflon tissues homogenizer). The homogenate were centrifuged at 10000 rpm for 30 minutes at 4°C ineffondorfe centrifuge. The supernatant obtained from the centrifugation were used for enzymes assay and protein estimation [2].
The supernatant of 10% tissues homogenates were used for the enzymes activity measurement. The enzymes AST, ALT, GGT and ALP activities were analysed employed by standard methods [8]. The tissues proteins were estimated by the Lowry method [9].
RNA was isolated from the selected rat tissues (liver, kidney and testis) using TRIzol reagent protocol were followed as manufacture guidelines. Briefly, 100 mg of fresh tissues were immediately frozen using liquid nitrogen homogenized in Teflon homogenizer in 1ml TRIzol reagent. Samples were centrifuged at 14,000 rpm for 10 min at 4°C, the supernatant transferred to new tube, 200ul chloroform were added and vertexes for 30sec, than centrifuged at 13,000 rpm for 15 min at 4°C . Aqueous layer transferred to new tube. RNA was precipitated by addition of the 500ul of isopropanol,samples were centrifuged at 13,000 rpm for 10 min at 4°C. The RNA pellet were obtained and washed with 70% Ethanol (three to four times). RNA pellet were resuspended in 100ul DEPC water. The RNA integrity were analysed by 2% (w/v) agarose gel electrophoresis. The purity and amount were measured in 260/280 nm absorption ratio for protein or phenol contamination. The cDNA was synthesised from isolated RNA employed with reverse transcriptase polymerase chain reaction (RT-PCR) by using AMV reverse transcriptase and total RNA (100 ng) from control and treated rat tissues used as a template. cDNA used template to amplified using gene specific primers such as, ALT Forward, 5’ CCGTGGAGAGGGTGGTGCCTTA 3’, Reverse, 3’ TGGGGTGCCGGGGTTGATG 5’ and AST Forward: 5’ GCGCCTCCATCAGTCTTTGC3’ Reverse: 3’ ATTCAT CTGTGCGGTACGCTC 5’ primer pairs. The ALT and AST enzymes PCR cycling profile are follows step 1: T 94°C for 10 min; step 2: T 94°C for 45 second; step 3: T 72°C for 30 second; step 4: T 72°C for 1 min; step 5: go to step 2 for 30 cycles and step 6: T 72°C for 10 min. These conditions, PCR were undertaken and yielded an exponential amplification. PCR amplified products were then separated by electrophoresis on a 2% (w/v) agarose gel and stained with ethidium bromide. The polynucleotide band intensities were measured by densitometry [10].
The experimental data were analyzed using Student’s t-test. Differences were considered significant when p<0.05.
Results
The ALT marker enzyme level increased for treated normal control and diabetic rat in heart, kidney and testis and in treated diabetic liver, whereas this enzyme activity reduced in the liver of treated normal control rat. The difference in the increase in ALT activity in these tissues ranged from 55-602% for treated diabetic group, whereas for the treated control group, the increase ranged between 11 and 338%. Normal control liver ALT however reduced 71% due to 28-HC treatment, and a further decrease in ALT activity was noted between diabetic control and normal control rat such as 38% reduction in diabetic heart, 91% in diabetic liver and 22% for diabetic testicular enzyme. In case of diabetic kidney enzyme, 67% increase in activity was noted against the control ALT activity (Table 1).
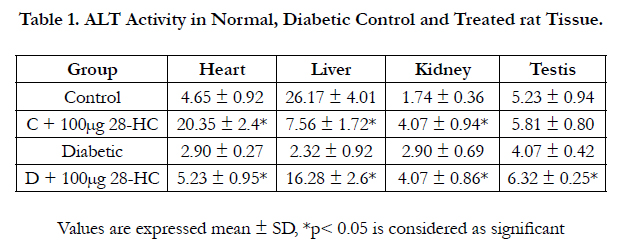

Table 1. ALT Activity in Normal, Diabetic Control and Treated rat Tissue.
In a similar manner, assessment of tissue AST enzyme activity yielded the following results. A 15% reduction in treated normal control liver and 50% reduction in the treated diabetic liver, 50% reduction in treated normal control testicular enzyme activity and 67% reduction in treated diabetic testicular enzyme activity. However, AST activity in treated kidney control raised 82% but decreased 67% in treated diabetic kidney. The rat heart AST activity increased 201% in the treated control tissue, but only 10% in treated diabetic heart tissue. Analysis of data between diabetic control and normal control AST activity indicated 25-234% increase in this enzyme activity in diabetic testis, kidney and heart, whereas a reduction was noted for the liver diabetic control to the extent of 38% (Table 2).
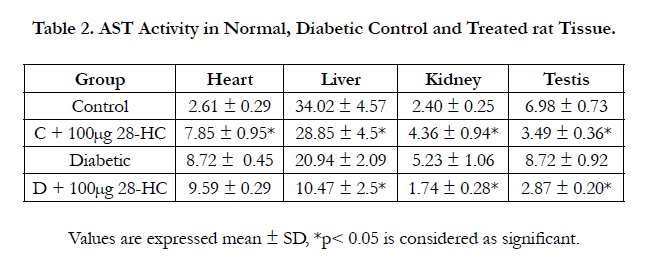
Table 2. AST Activity in Normal, Diabetic Control and Treated rat Tissue.
The gama glutamyl transpeptidase activity increased 15% in the liver of the treated control and 50% in the liver of treated diabetic rat for the same amount (100μg) of 28-HC administered by oral gavge for the said duration. The diabetic control rat yielded 25% less of this enzyme activity compared to the normal control. The heart GGT activity was raised 75% in the treated normal control and 62% in the treated diabetic animal with only 12% increase in diabetic control versus normal control. Rat kidney GGT enzyme activity was elevated 125% in diabetic control whereas this enzyme activity reduced 83% in the diabetic animal following 28-HC administration. However kidney GGT activity increased 226% in the treated control rat. In a similar manner the testicular GGT activity of the treated diabetic rat decreased 37% whereas the diabetic control exhibited an activity increase of 253% while the treated normal control exhibited an increase of 57% (Table 3). The alkaline phosphatase (ALP) activity as a liver marker enzyme was determined in a similar manner following treatment of normal and diabetic rat with 28-HC by oral gavage administration for a duration as reported early. The liver enzyme activity was noted as elevated for the diabetic control (67%), for the treated diabetic rat (133%) and for the treated normal control (167%) (Table 4).
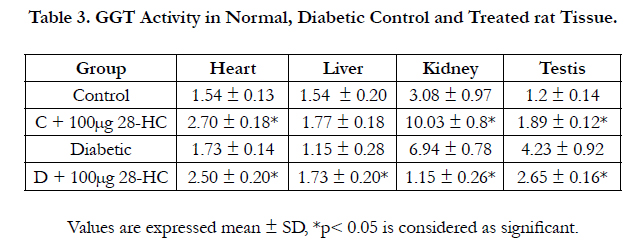
Table 3. GGT Activity in Normal, Diabetic Control and Treated rat Tissue.
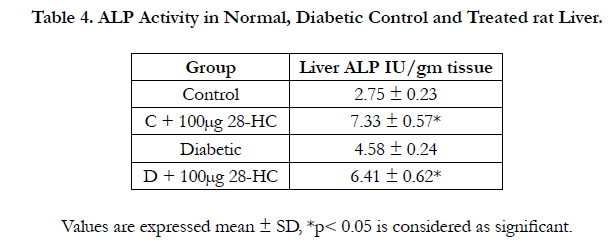
Table 4. ALP Activity in Normal, Diabetic Control and Treated rat Liver.
Gene expression studies were carried out on a set of selected marker genes employing specific tissue mRNA, generating corresponding cDNA using AMV reverse transcriptase and amplified each cDNA population by the polymerase chain reaction (PCR) standardized for use of the pair of the specific primers that ranged between 19-24 mers. The specific nucleotide sequence of each pair of primers is as in under methods. Following PCR amplification the ethidium bromide band intensity of the amplicons were assessed using a densitometric software in a gel documentation system.
Figure 1. In case of liver, ALT an 180bp band was noted in the control and test lanes. For the AST, the band size was 280bp in both lanes. The 28-HC treated group’s ALT band intensities were noted as 80 fold greater for treated normal control and 60 fold higher for treated diabetic rat than that for the respective control bands. ALT gene expression in kidney yielded similar band sizes whereas difference in the band intensity for the treated ALT bands was only 50 fold higher than the respective control. Using the same pair of primers, the ALT gene amplicons of testicular tissue yielded intensity difference of 60 fold between test and control bands of ALT gene expression. The 28-HC treated rat liver tissue AST band intensities were noted as 80 fold greater for treated normal control and 60 fold higher for treated diabetic rat than that for the respective control bands. AST gene expression in kidney reduced the band intensity for the normal treated AST band was only 50 fold than the normal control and 60 fold in diabetic treated. Using the same pair of primers, the AST gene amplicons of testicular tissue yielded 60 fold in diabetic treated compared to diabetic control gene expression.
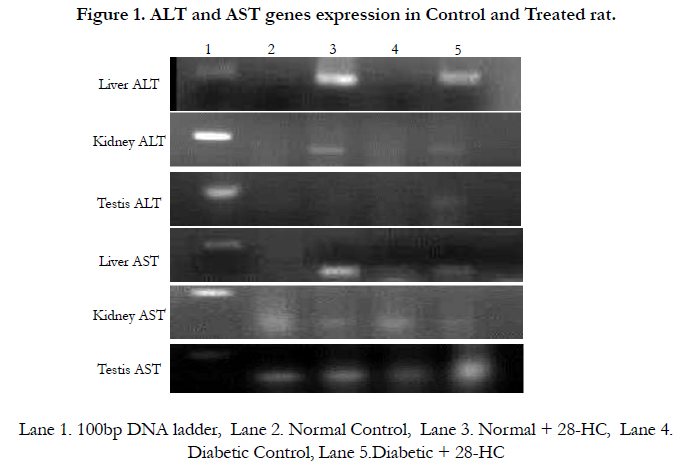
Figure 1. ALT and AST genes expression in Control and Treated rat.
Discussion
Enzymes levels in the various tissues studied earlier indicated significantly increased ALT activity in treated control and diabetic liver. Increase in ALT enzyme activity suggests increase transamination of the amino acid alanine to α-ketoglutarate, a reactive intermediate in the crabs TCA cycle leading to the formation of pyruvate [4]. Elevated pyruvate levels in a cell contribute to bio energy production through the formation of ATP [5]. Hence it can be safely presume that 28-HC administration can causes ATP synthesis in the cells of tissues. However the reduced ALT activity in treated control liver begs a reason for the observed reduction, so was the case in diabetic heart, diabetic liver and diabetic control rat testis, However down ward changes in ALT activity between normal and diabetic rat suggest that bio energetics and in diabetic tissues exist negative to that in normal tissue [11]. With the exception with diabetic kidney that exhibited a significant positive change in this enzyme activity wises the control. The major exception was that of normal control ALT whose activity down regulated around 70% by 28-HC below that of untreated control, this is raises the question as to whether the ALT activity lowering effect shown by 28-HC is beneficial for the liver tissue. Assessment of the AST and ALT ratio for this tissue should reveal the truth [12]. Assessment of the AST enzyme activity indicated significant down ward trend in this enzyme activity in treated normal control liver and testis and in treated diabetic liver and diabetic testis suggesting reduced transamination between aspartate and glutamate contributing to the reduced generation of alpha-ketoglutarate and oxaloacetate [6]. AST/ALT ratio greater than 2:1 is indicative of liver injures and off hepatotoxicity [13]. The concomitant increase in Gamma-glutamyl transferase (gamma-glutamyl transpeptidase) enzyme activity in treated control and diabetic rat liver as observed in this study is further confirmation for hepatotoxicity. The AST/ALT ratio of control liver was estimated to be 0.1 that of treated control liver was 1.04. The diabetic control liver AST/ALT ratio was found to be 9.03, indicative of severe liver injury. However, 28-HC treated diabetic liver AST/ALT ratio registered a value of 0.643 suggestive of probable renormalization of the liver injury caused due to diabetes.
Gamma glutamate transpeptidase (transferase) (GGT) catalysis the transfer of gamma glutamyl functional group to an amino acid, peptide, water (forming glutamate) it’s a significant marker enzyme for the liver in xenobiotic metabolism [14]. Elevated GGT activity suggests disease of the liver, biliary system and pancreases. It is also considered to correlate with CVD and atherosclerotic plaque [15]. Alongside ALP enzyme activity also measured as biliary marker, alcoholic liver disease. Although specifically serum GGT enzyme activity was not estimated, tissue GGT activity was measured before and after 28-HC administration it was found that the tissue GGT generally indicated significant elevation following 28-HC treatment. Use of 28-HC however reduced kidney GGT activity following treatment of the diabetic animal. Reduction in GGT activity was also seen in treated diabetic testis. Elevation in tissue GGT activity can translate to an elevation in serum GGT levels suggestive of CVD or atherosclerotic plaque in the treated rat. Since assessment of cholesterol and triglyceride levels noted reduction in LDL, VLDL levels the probability of developing atherosclerotic plaque [2]. The raised GGT level in the tissues should therefore be consider as due to the induction of GGT production as a consequence of 28-HC effects at the genetic level. Therefore any observed increases in serum GGT need not necessarily reflect cardio vascular disease (CVD). The marker status of this enzyme and other enzyme studied therefore is influenced by the dietary amounts of 28-HC and constituents that are capable of gene induction at the tissue level. Serum levels of marker enzymes are generally considered as these which have been synthesised in the tissue and released in to the blood circulation. Hence, based on observation made in this study dependence on serum marker status for clinical diagnosis become questionable. Reduction in tissue GGT activity in rat liver of the diabetic control rat and in rat kidney following administration of 28-HC of is suggestive of suppression of GGT gene expression in these tissues due to diabetes on the one hand and due to 28-HC on the other. A similar situation is also noted in treated diabetic rat testis. The differential expression of GGT activity in the various tissues, control verses treated, indicates the differential effects in gene expression in diabetic and control rat tissues as well as due to the effect of 28-HC administration.
In case of the tissue alkaline phosphatase, the liver enzyme activity was augmented in the diabetic rat as well as in treated control and diabetic rats this observation further confirms the tissue related responses due to diabetes and due to the effect of the dietary constituents. Differential yield as a tissue response to diabetes and 28-HC was again noted with respect to the tissue status of marker [16].
Conclusion
Dietary plant keto steroid, 28-homocastasterone enhances the transaminase and phosphatase enzymes catalytic activity in the tissues differentially, resulted to cellular transamination and phosphorylation status affected by this plant steroid. It is suggested that 28-HC supplemented diets may be suitable for higher energy related work activities in man and animals.
Acknowledgement
The authors acknowledge Dr.V.S.Pori, National Chemical Laboratory (CSIR), Pune, India, for her gift of 28-homocastasterone used in this study. Author Mr.V.Athithan gratefully acknowledges University Grant Commission (UGC), New Delhi, India for financial support received as Research Fellowship.
References
- Premalatha R, Srikumar K, Vijayalaksmi D, Kumar GN, Mathur PP (2014) 28-Homobrassinolide: a novel oxysterol transactivating LXR gene expression. Molecular biology reports 41(11): 7447-7461.
- Athithan V, Premalatha R, Srikumar K (2014) Down Regulation of Plasma and Tissue Biomarkers by Homocastasterone. International Journal of Drug Delivery 6(2): 172-178.
- Athithan V, Premalatha R, Mukherjee V, Mani N, Srikumar K (2015) HOMOCASTASTERONE: A NOVEL PLANT KETOSTEROID INDUCING HAEMATOLOGICAL CHANGES IN NORMAL AND DIABETIC MALE RAT. International Journal of Pharmacy and Pharmaceutical Sciences 7(7): 1-4.
- Ozer J, Ratner M, Shaw M, Bailey W, Schomaker S (2008) The current state of serum biomarkers of hepatotoxicity. Toxicology 20245(3): 194-205.
- Hertz L, Peng L, Dienel GA (2007) Energy metabolism in astrocytes: high rate of oxidative metabolism and spatiotemporal dependence on glycolysis/ glycogenolysis. Journal of Cerebral Blood Flow & Metabolism 27(2): 219-249.
- Weinberg F, Hamanaka R, Wheaton WW, Weinberg S, Joseph J, et al., (2010) Mitochondrial metabolism and ROS generation are essential for Kras-mediated tumorigenicity. Proceedings of the National Academy of Sciences. 107(19): 8788-8793.
- Kumar GN, Lakshmy S, Srikumar K (2010) Negative modulation of alkaline phosphatase and creatine kinase by homobrassinolide. International Journal of Drug Delivery 2(3): 258-264.
- Reitman S, Frankel S (1957) A colorimetric determination of oxaloacetic and glutamic pyruvic transaminases. American journal of clinical pathology 28(1): 56 – 63.
- Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) Protein measurement with the Folin phenol reagent. J biol Chem 193(1): 265-275.
- Maniatis T, Fritsch EF, Sambrook J (1982) Molecular cloning: a laboratory manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory. 5(1): 104.
- Sikaris KA (2004) The clinical biochemistry of obesity. The Clinical Biochemist Reviews. 25(3): 165-181.
- Nyblom H, Berggren U, Balldin J, Olsson R (2004) High AST/ALT ratio may indicate advanced alcoholic liver disease rather than heavy drinking. Alcohol and alcoholism 39(4): 336-339.
- Giannini EG, Testa R, Savarino V (2005) Liver enzyme alteration: a guide for clinicians. Canadian medical association journal 172(3): 367-379.
- Zhang H, Forman HJ, Choi J (2005) γ‐Glutamyl Transpeptidase in Glutathione Biosynthesis. Methods in enzymology 401: 468-483.
- Lum G, Gambino SR (1972) Serum gamma-glutamyl transpeptidase activity as an indicator of disease of liver, pancreas, or bone. Clinical Chemistry 18(4): 358-362.
- Lum G (1995) Significance of low serum alkaline phosphatase activity in a predominantly adult male population. Clinical chemistry 41(4): 515-518.





