Role of Acid-sensing Ion Channels (ASICs) in the Vascular Endothelial Cells Injury of Henoch- Schonlein Purpura Children
Li-ping Yuan1*, Qi-di Peng1, Yan Bo2, Guo Xiao-yan1, Hu Bo1
1 Department of Pediatrics, First Affiliated Hospital of Anhui Medical University, Hefei city, Anhui province, China.
2 Department of Medical Technology, Anhui Medical College, Hefei city, Anhui Province, China.
*Corresponding Author
Li-ping Yuan,
Department of Pediatrics, The First Affiliated Hospital of Anhui Medical University,
Hefei, 230022 China.
Tel: (+ 86)-551-6292-2058
E-mail: yuanliping3986@sina.com
Received: June 12, 2016; Accepted: August 04, 2016; Published: August 08, 2016
Citation: Li-ping Yuan, Qi-di Peng, Yan Bo, Guo Xiao-Yan, Hu Bo (2016) Role of Acid-sensing Ion Channels (ASICs) in the Vascular Endothelial Cells Injury of Henoch-Schonlein Purpura Children. Int J Clin Med Allergy. 4(2), 44-51. doi: dx.doi.org/10.19070/2332-2799-1600010
Copyright: Li-ping Yua© 2016. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
Objective: Acid-sensing ion channels (ASICs) are cationic channels that are activated by extracellular acidification. ASICs are widely expressed in the peripheral and central nervous systems and participate in many important physiological and pathological processes, such as synaptic plasticity, cerebral ischemia, and pain. Recently accumulating evidence suggests that ASICs can also profoundly affect the physiological properties of non-neural cells. This study was firstly to investigate the expression of ASICs subunits in the vascular endothelial cells of Henoch-Schönlein Purpura (HSP) patients and their role in vascular endothelial cell injury following acid exposure.
Methods: Human dermal microvascular endothelial cells (HDMECs) were used to establish the model of vascular endothelial cell injury by incubation with IgA1 from HSP patients. The cell injury following acid exposure was analyzed with inflammatory cytokines release assay and cytoskeletal protein destrin, α-skeletal muscle actin (SM-α) changes. The acid- induced ASICs, destrin and SM-α actin mRNA and protein expressions in HDMECs were evaluated by quantitative real-time polymerase chain reaction (qPCR) and western blotting, respectively. The release of inflammatory cytokines interleukin-8 (IL-8), thrombomodulin (TM) and nitric oxide (NO) were analyzed by ELISA methods. Furthermore, we analyzed the relationship between ASICs expression with cytokines release, destrin and SM-α actin levels by Person Correlations.
Results: The results showed that extracellular acid could upregulate ASIC1a, ASIC2a and ASIC3 mRNA and protein expression, stimulate IL-8, TM and NO, and decrease destrin and SM -α actin expression in HDMECs pretreated with IgA1 from HSP patients. Furthermore, ASICs expression was positively related to the production of cytokines and negatively related with destrin and SM-α actin expression, respectively. In addition, pharmacological blockade of ASICs attenuated the inflammation response and inhibited the loss of cytoskeletal protein in vascular endothelial cells pretreated with IgA1 from HSP patients.
Conclusion: These findings showed that extracellular acid could activate ASICs expression in the vascular endothelial cells pretreated with IgA1 from HSP patients, which induced further endothelial cells damage. Additionally, the inhibitor of ASICs may have a significant protective effect on the inflammatory injury of vascular endothelial cells.
2.Background
3.Materials and Methods
3.1.Patients and controls
3.2.Isolation of IgA1
3.3.Culture of Human dermal microvascular endothelial cells (HDMECs)
3.4.Experimental design
3.5.Measurement of interleukin-8 (IL-8), thrombomodulin(TM) and nitric oxide (NO) endogenous
3.6.Analysis of ASICs and destrin, α-skeletal muscle (SM-α) actin mRNA expression
3.7.Western blot analysis
3.8.Statistical analysis
4.Results
4.1.Effects of extracellular acidification on the expression of ASICs mRNA and protein in cultured HDMECs
4.2.Effects of extracellular acidification on IL-8, TM and NO release in the supernatant of HDMECs
4.3.Effects of extracellular acidification and on destrin, SM-α actin expression in HDMECs
5.Discussion
6.Conclusion
7.Acknowledgement
8.References
Keywords
Acid-Sensing Ion Channels (ASICs); Henoch-Schonlein Purpura (HSP); Vascular Endothelium Cells.
Background
Henoch-Schönlein Purpura (HSP) is a common small vessel vasculitis in children which is characterized by non-thrombocytopenic purpura, arthritis, bowel angina and glomerulonephritis [1]. It is generally accepted that immunoglobulin A (IgA), especially IgA1 and complement C3 deposited in the walls of arterioles, capillaries, and venules, immune inflammatory reaction are involved in the vascular endothelial cell injury of HSP patients [1-3]. In the majority of cases, HSP is a self-limiting disease and treatment is supportive. However, HSP has a high rate of recurrence, and some patients can progress to HSP nephritis (HSPN), which can result in renal failure [4]. Up to now, there is no effective therapy to impede these events to occur. So it is extremely urgent to further research on the pathophysiology of HSP.
Acid-sensing ion channels (ASICs) are cationic channels which belong to the degenerin/ epithelial Na+ channel (DEG/ENaC) superfamily and activated by extracellular protons [5]. To date, at least six subunits of ASICs have been identified: 1a, 1b, 2a, 2b, 3, and 4 [6]. ASIC1a is the only subunit that is permeable to Ca2+, as it differs from other ASIC subunits. Recent studies have indicated that ASIC1a is involved in synaptic plasticity, learning, and memory [2, 3]. Activation or sensitization of Ca2+-permeable ASIC1a has also been demonstrated to be responsible for acidosis-mediated ischemic brain injury caused by Ca2+ influx in neurons [4].
In HSP patients, the inflammatory damage is characterized by the presence of leukocytes in the vessel and the IgA1-IC deposition and then the swelling and damage of vessel mural structures. The lumen is narrowed or occluded leading to ischemia and necrosis [11], which lead to the decrease of pH in the local tissue. The acidosis in the local tissue would deteriorate the injured vascular endothelial cells of HSP. So identification of how the sensors of vascular endothelial cells adjust to these pH changes is of primary importance to understanding the processes of underlying the injured vascular pathologies associated with HSP. Intriguingly, our recent study found that ASIC1a and ASIC3 expression significantly increased in the vascular endothelium of HSP patients than those of normal healthy children [12]. But the role of ASICs in the acid-induced vascular endothelial cell injury in HSP patients has not been elucidated. This study examined the expression of different ASIC subunits in vascular endothelial cells when extracellular acidosis occurred and investigated the role of ASICs in vascular endothelial cell injury of HSP patients.
Materials and Methods
10 children (5 boys and 5 girls, range from 3 to 14 years old) with acute onset and/or active presentation of HSP were enrolled in this study from the first affiliated hospital of Anhui Medical University during January 2015 to June 2015. The diagnosis of HSP was based on the criteria defined by the European League Against Rheumatism, Pediatric Rheumatology International Trials Organization, and Pediatric Rheumatology European Society (UULAR/PRINTO/PRES) 2010 [13].10 healthy subjects (5 boys and 5 girls, range from 3 to 14 yeas old) were recruited as normal controls. The approval and fully informed counseled consent were obtained from the ethical committee of the first affiliated hospital of Anhui Medical University, Anhui Medical University and the children's parents, respectively.
5ml sera were isolated from peripheral blood of HSP and healthy children following standard protocols. IgA1 was isolated from sera by Immobilized Jacalin (Pierce, Rockford, IL, USA), a commercial kit that can isolate serum-circulating IgA1 by galactosebinding lectin. Because the amount of polymeric IgA1 (pIgA1) recovered from the purification process was not sufficient for further analyses, we incubated the purified monomeric IgA1 (mIgA1) at 63°C for 150 min to obtain aggregated IgA1 (aIgA1) as described previously [14, 15]. The transition from mIgA1 to aIgA1 was monitored using a Sephacryl S-200 column (GE, Centennial Avenue Piscataway, NJ USA), and a single peak was observed after incubation at 63°C. The purified IgA1 was identified by western blotting and stored at -70°C for further analysis.
HDMECs were obtained from Jennio Biotechnology limited company of GuangZhou and were cultured at 37°C in 5%CO2 humidified incubator with RPMI-1640 medium containing 15% fetal bovine serum. When the cells growth became confluent two days later, they were used to do the following experiments.
HDMECs were plated in 6-well plates at 1×106 cells/well and pretreated with aIgA1 (final concentration was 250ug/ml) from HSP patients or healthy controls for 6h according to the previous study reported [16]. The supernatants were removed and pH6.5 solution was added into the cells for 4h. The cells were divided into these groups: 1.blank control group (pH7.4 solution without aIgA1); 2. pH6.5 (pH6.5 solution without aIgA1); 3. normal IgA1+pH6.5(pH6.5 solution with aIgA1 from normal healthy children); 4. normal IgA1(pH7.4 solution with aIgA1 from normal healthy children); 5. HSP IgA1(pH7.4 solution with aIgA1 from HSP patients); 6. HSP IgA1+pH6.5(pH6.5 solution with aIgA1 from HSP patients). Each well was then washed by PBS for two times.
To investigate the role of ASICs in vascular endothelial cell injury of HSP patients, the cells were also treated with final concentrations of Amiloride of 100μmol/L or PcTX1 of 20nmol/L. After 30 min, the cells were treated with pH 6.5 solutions for 4h; the pH and time was determined in a pilot study. After treatment, the cells continued in normal media for about 4h. We did not see a change in cell viability following pH treatment as measured by trypan blue dye exclusion assay. And the supernatants were collected and particulate material removed by centrifugation for 10 min at 1000g. The cells were also collected at the same time. All the experiments were performed in triplicate.
The concentration of IL-8, TM and NO in the supernatants of HDMECs was determined by commercial ELISA kits, according to the manufacturer’s instructions (Yuanye, Shanghai, China). The absorbance was measured with a microplate reader (Labsystems Multiskan Bichromatic).
Total RNA was extracted from HDMECs using TRL reagent (OMEGA, USA) according to the manufacturer’s instructions (OMEGA, USA). The concentration and purity of RNA were determined spectrophotometrically at ratio 260/280, respectively. 1ug RNA was used for single-strand cDNA synthesis with the Revert AidTM First Strand cDNA Synthesis system (Thermo, USA) for qPCR kits. Primers (Sangon,Shanghai, China) used for quantitative real-time polymerase chain reaction (qPCR) analysis of ASIC1a, ASIC2a, ASIC3, destrin, SM-α actin and β-actin were listed on Table 1. The mRNA levels of ASICs and destrin, SM-α actin were determined by quantitative realtime polymerase chain reaction (qPCR). The RT product was used for PCR in a 25µl reaction volume containing 12.5ul Maxima SYBR Green/ROX qPCR Master Mix (Thermo, USA) and 0.25 uM each of forward nd reverse primers. The qPCR was under the following conditions: started at 50° for 2 min and 95° for 10min (initial denaturation), followed by a 40-cycle amplification (denaturation at 95°for 15 s, annealing/extension at 60° for 60s). The mRNA levels of these ASICs and destrin, SM-α actin were normalized with those of β-actin mRNA.
The protein concentration of lysates was determined by BCA Protein Assay Kit (Beyotime, Shanghai, China). Proteins (20ug) were separated using 10% (13% for caspase analysis) SDS-PAGE and transferred to poly-vinylidene fluoride membranes. The membranes were probed with primary affinity-purified rabbit polyclonal antibody against ASIC1a , ASIC2a, ASIC3 and destrin, SM-α actin (Abcam, HongKong, England) followed by secondary goat anti-rabbit alkaline-phosphatase-conjugated antibody (Promega). The blots were washed with TBST, and then developed by enhanced chemiluminescence detection regents (ECL, Amersham). The protein bands were quantified by the average ratios of integral optic density following normalization to the housekeeping gene.
All tests were performed using SPSS 16.0 software. All results were expressed as mean ± SD, Statistical differences between groups were determined according to one-way analysis of variance (ANOVA), unpaired student t-test, person correlations. P<0.05 was considered significant. The relationship of ASICs expression with cytokines, destrin, SM-α actin was analyzed by Person Correlations.
To evaluate the expression of ASIC subunits in the vascular endothelial cells of HSP, we isolated IgA1 from HSP patients to establish the model of injured vascular endothelial cells of HSP and found that ASIC1a, ASIC2a and ASIC3 mRNA and protein expression in HDMECs were higher in group 5 (HSP IgA1) than those in group 4 (normal IgA1) and group 1 (blank control). This result suggested that ASICs expression was upregated in vascular endothelial cells pretreated with IgA1 from HSP patients.
Moreover our results showed that extracellular acidosis alone could upregulate ASIC1a, ASIC2a, ASIC3 mRNA and protein expression of HDMECs. There was significant difference between group 2 (pH6.5) and group 4 (normal IgA1), group 2 (pH6.5) and group 5 (HSP IgA1) (P<0.01). In addition, ASIC1a, ASIC2a, ASIC3 mRNA and protein levels of HDMECs in group 6 (HSP IgA1+pH6.5) were much higher than those in other groups (P<0.01 Figure 1 A, B, C), which suggested that extracellular acidosis could further increase ASICs mRNA and protein levels in vascular endothelial cells pretreated with IgA1 from HSP patients.
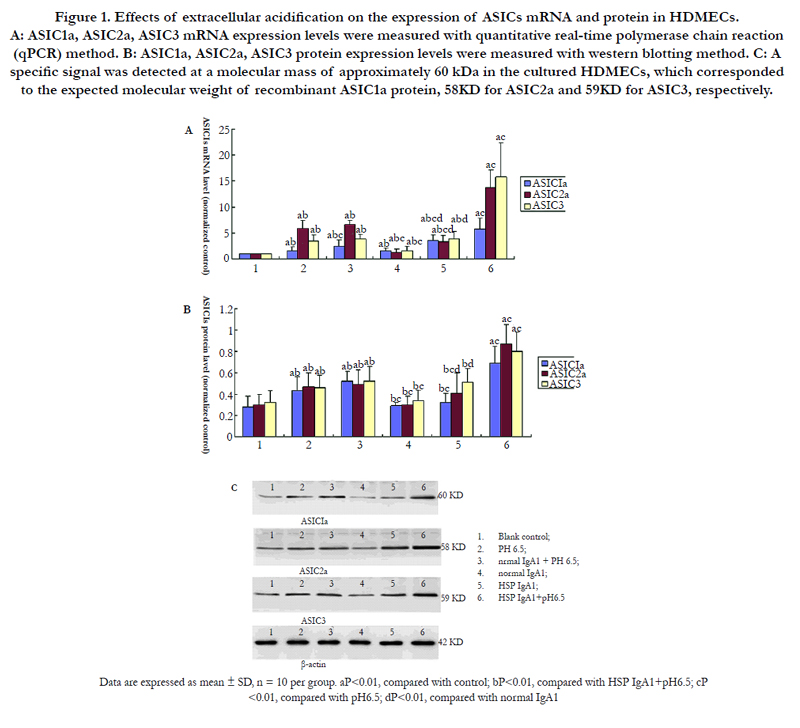
Figure 1. Effects of extracellular acidification on the expression of ASICs mRNA and protein in HDMECs. A: ASIC1a, ASIC2a, ASIC3 mRNA expression levels were measured with quantitative real-time polymerase chain reaction (qPCR) method. B: ASIC1a, ASIC2a, ASIC3 protein expression levels were measured with western blotting method. C: A specific signal was detected at a molecular mass of approximately 60 kDa in the cultured HDMECs, which corresponded to the expected molecular weight of recombinant ASIC1a protein, 58KD for ASIC2a and 59KD for ASIC3, respectively.
It is well-known that increased inflammatory cytokines were associated with the inflammatory injury of vascular endothelial cells of HSP patients [2]. Cell injury was assayed by the measurement of IL-8, NO and TM release in HDMECs. Figure 2 showed that IL-8, NO and TM levels were higher in the supernatant of HDMECs in group 10 (HSP IgA1) than those in group 9 (normal IgA1) and group 1 (blank control). These results suggested that the injured vascular endothelial cells of HSP patients could secret some cytokines, such as IL-8, NO and TM. IL-8, NO and TM levels in the supernatant of HDMECs were much higher in group 6 (HSP IgA1+pH6.5) than those in other groups when these cells are exposed to pH6.5 solutions, which showed that extracellular acidosis could further the vascular endothelial cell injury of HSP patients.
To determine whether activation of ASICs is involved in acid-induced vascular cell injury in HSP patients, we tested the effect of ASICs blockers Amiloride or PcTX1 on inflammatory cytokines release. As shown in Figure 2, at pH6.5 vascular endothelial cells incubated with aIgA1 from HSP patients and normal healthy children induced substantial IL-8, NO and TM release that was reduced by Amiloride or PcTX1 (P<0.01). These results suggested that ASICs might be involved in the development of acid-induced vascular endothelial cells damage of HSP patients.
In addition, we analyzed the relationship of ASICs with inflammatory cytokines. Our study showed that IL-8, TM and NO release in HDMECs pretreated with aIgA1 from HSP patients were positively related to the level of ASIC1a, ASIC2a and ASIC3 mRNA expression, as demonstrated in Figure 3.
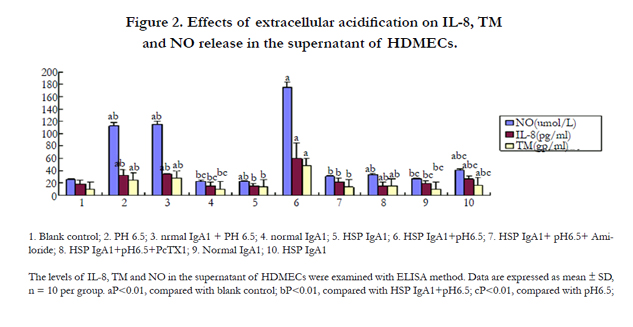
Figure 2. Effects of extracellular acidification on IL-8, TM and NO release in the supernatant of HDMECs.
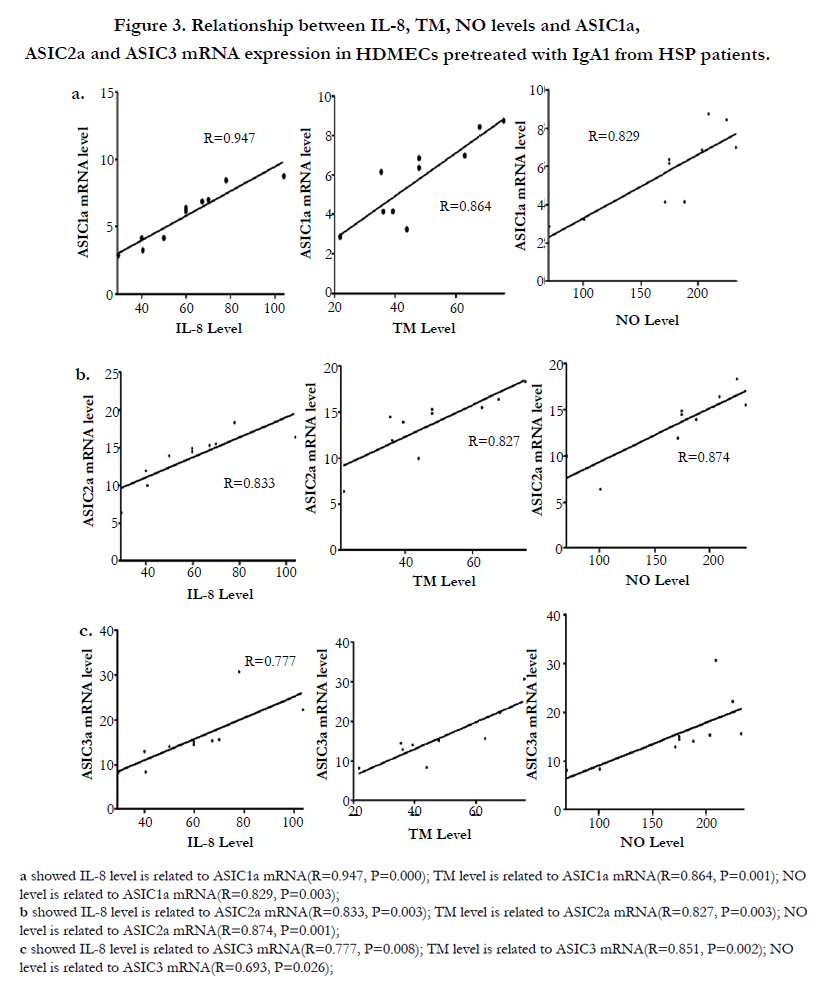

Figure 3. Relationship between IL-8, TM, NO levels and ASIC1a, ASIC2a and ASIC3 mRNA expression in HDMECs pretreated with IgA1 from HSP patients.
Destrin and SM-α actin are structure proteins of vascular smooth cells (VSMCs) which are crucial in cell motility, movement and contraction. The results showed that extracellular acid damaged destrin and SM-α actin mRNA and protein expression in vascular endothelial cells. Destrin and SM-α actin mRNA and protein expression was obviously decreased in HDMECs incubated with IgA1 from HSP patients and normal healthy children in low pH solution, and there was significant difference between them (P<0.01, P<0.01) (Figure 4A, Figure 4B).
To further investigate whether the enhanced ASIC expression in low pH environment was involved in the destruction of blood vessel of HSP patients, we examined destrin and SM-α actin expression in vascular endothelial cells by blocking ASICs with Amiloride or PcTX1. Figure 4 showed that decreased destrin and SM-α mRNA and protein expression in HDMECs incubated with IgA1 from HSP was abolished by Amiloride or PcTX1 (P<0.01) as well as in HDMECs incubated with IgA1 from normal healthy children (P<0.01).
Moreover Figure 5 demonstrated that destrin and SM-α mRNA level is negatively related to the mRNA expression of ASIC1a (R=-0.713, P=0.021; R=-0.867, P=0.001), ASIC2a (R=-0.842, P=0.002; R=-0.798,P=0.006) and ASIC3 (R=-0.671, P=0.034; R=-0.742,P=0.014) in HDMECs incubated with IgA1 from HSP patients.
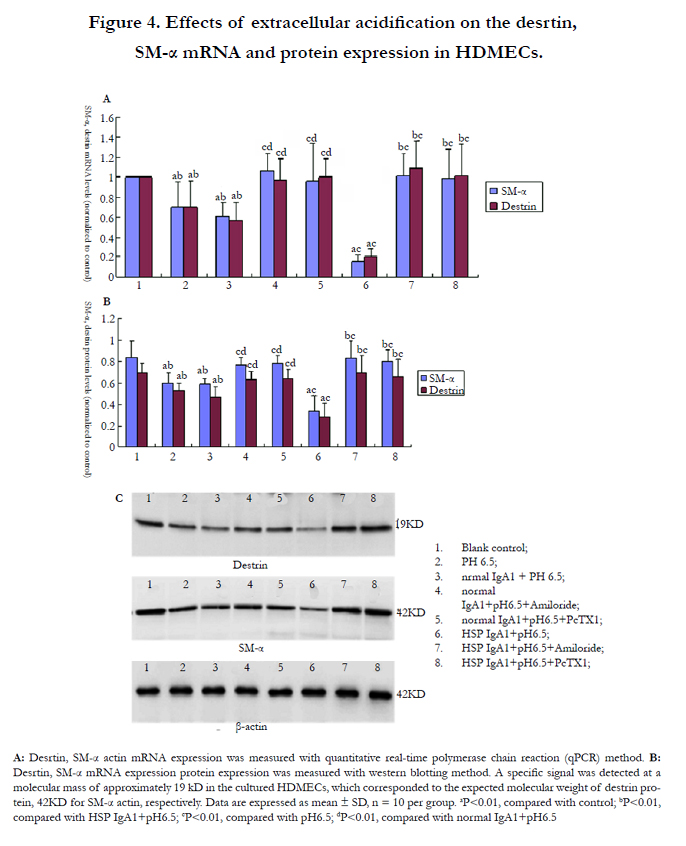
Figure 4. Effects of extracellular acidification on the desrtin, SM-α mRNA and protein expression in HDMECs.
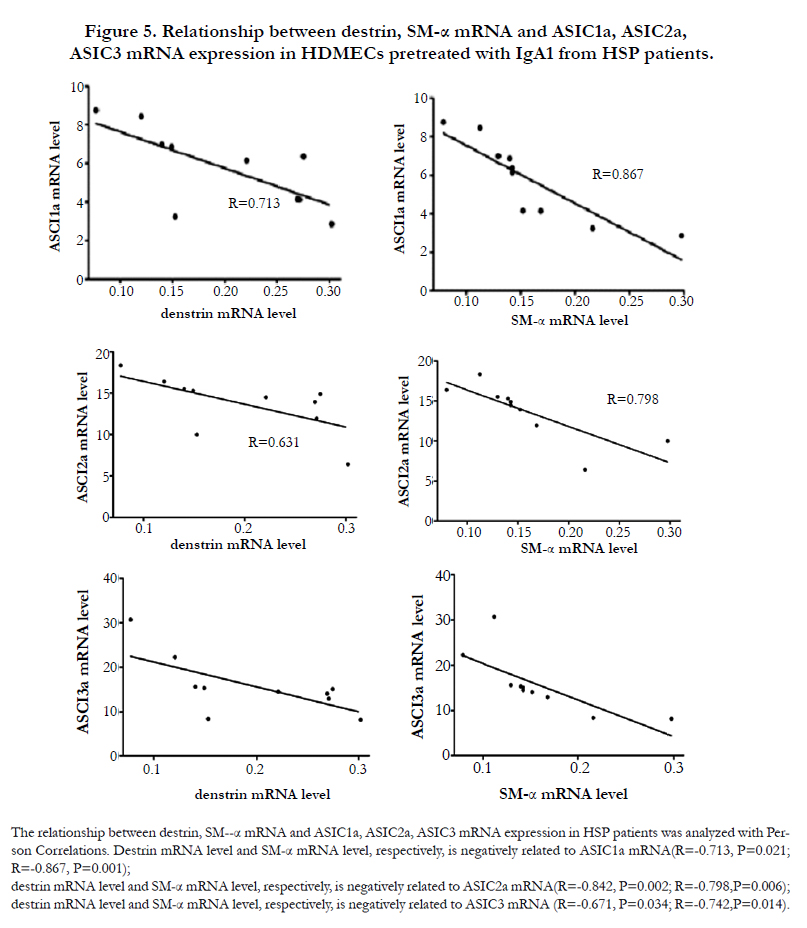
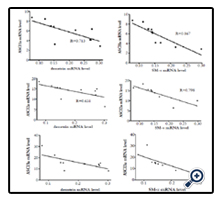
Figure 5. Relationship between destrin, SM-α mRNA and ASIC1a, ASIC2a, ASIC3 mRNA expression in HDMECs pretreated with IgA1 from HSP patients.
Discussion
ASICs are members of the DEG/ENaC superfamily and expressed mainly in the mammalian nervous system and other tissues or cells such as the brain, bone, taste receptors, carotid body, glioma cells, disc cells and vascular smooth muscle cells. Our recent study demonstrated ASIC1a, ASIC3 protein also exited in the skin vessel endothelium of HSP patients and ASIC1a, ASIC3 expression was upregulated by acute sera from HSP patients in human umbilical vein endothelial cells [12]. In this study, we examined whether ASICs subunits were expressed in vascular endothelial cells that were exposed to acidic solutions under inflammatory environmental conditions. qPCR and Western blotting indicated increased ASIC1a, ASIC2a and ASIC3 mRNA and protein expression in vascular endothelial cells, especially in the cells pretreated with IgA1 from HSP patients. The pH value in the inflammation area is always lower than that in normal tissues, ranging from pH 5.5 to 7, while pH50 values of 6.2-6.7 have been measured for ASIC1a and ASIC2a and ASIC3 with a pH50 of 4.1-5.0 [17]. Our pilot study found that ASIC1a and ASIC2a expression reached the maximum when pH lowered to pH6.5. These results indicated that functional ASIC subunits were highly expressed in vascular endothelial cells of HSP. However the function of ASICs in vascular endothelial cell injury of HSP patients has not yet been identified.
Many research revealed that circulating IgA1 (especially aberrant O-linked glycosylation of immunoglobulin A1) in sera of children with active HSP separated and purified by Immobilized Jacalin could bind to vascular endothelial cells and induce endothelial cell inflammatory injury. During the inflammatory state in HSP-patients, the extracellular pH decreases because of cell damage, immune response, or hypoxic metabolism; then, acid-sensitive channels are activated and perceive the acidic microenvironment, contributing to endothelial cells damage or further these cells damage. In this study, we isolated IgA1 from HSP patients to establish the model of vascular endothelial cell injury of HSP patients as previously described and investigated the role of ASICs in acid-induced vascular endothelial cell injury of HSP patients. The cell injure following acid exposure was analyzed with measurement of inflammatory cytokines release and cytoskeleton protein changes.
It has been showed that sera (or IgA1 isolated from sera) derived from acute stage of HSP had the ability to enhance IL-8 production by human umbilical venous endothelial cells [16]. Thrombomodulin (TM) is a membrane-bound glycoprotein predominantly located at the vascular endothelium, which is released from the surfaces of endothelial cells when endothelial was injured [17]. NO is proved to be a secondary effect after the interaction of eNOS with Ca2+/cal modul in increased levels of and NO in endothelial cells produce proinflammatory profiles, which might lead to vascular diseases [18]. Our study showed that extracellular acid could induce the release of IL-8, TM and NO in vascular endothelial cells pretreated with aIgA1 from HSP patients, which was positively correlated with ASIC1a, ASIC2a and ASIC3 expression, respectively. In addition, ASIC blockers, Amiloride and PcTx1, inhibited cytokines release induced by extracellular acid. These results indicated that H+-evoked ASICs activation contributed to cell injury in vascular endothelial cells or further the cells injury.
Destrin is responsible for the rapid rearrangement of actin structures in the cells [19], and has been shown to play a crucial role in cell motility [20]. α-skeletal muscle (SM-α) actin is a kind of structure protein of vascular smooth muscle cells (VSMCs) which is crucial in cell movement and contraction [21]. Studies reported that the expression of SM-α decreased significantly in VSMCs due to some damage factors caused VSMCs damage [22]. In this experiment we also found that the destrin and SM-α expression was reduced by extracellular acid in vascular endothelial cells pretreated with IgA1 from HSP patients while Amiloride and PcTx1 could abolish the effect. Moreover destrin and SM-α expression was negatively related to ASIC1a, ASIC2a and ASIC3 expression. This was consistent with Grifoni et al reported [23].
Conclusion
The present study showed that ASIC1a, ASIC2a and ASIC3 was present in vascular endothelial cells and was highly upregulated in vascular endothelial cells pretrested with IgA1 from HSP patients with a reduction in the extracellular pH. These findings also provided evidence that ASICs was involved in vascular endothelial cells injury processing and the inhibitors of ASICs may represent candidates for the therapy of HSP patients.
Acknowledgements
This research was supported by the Project supported by the National Natural Science Foundation of China (NO 81471617) and Key University Science Research Project of Anhui Province (NO KJ2015A360).
References
- Yang YH, Chuang YH, Wang LC, Huang HY, Gershwin ME, et al., (2008) The immunobiology of Henoch-Schonlein purpura. Autoimmunity Reviews.7(3): 179-184.
- Besbas N, Saatci U, Ruacan S, Ozen S, Sungur A, et al., (1997) The role of cytokines in Henoch Schonlein purpura. Scand J Rheumatol 26(6): 456- 460.
- Tsuji Y, Abe Y, Hisano M, Sakai T (2004) Urinary leukotriene E4 in Henoch- Schonlein purpura. Clin Exp Allergy. 34(8): 1259-1261.
- Ronkainen J, Nuutinen M, Koskimies O (2002) The adult kidney 24 years after childhood Henoch-Schonlein purpura: a retrospective cohort study. Lancet 360: 666-670.
- Waldmann R, Champigny G, Bassilana F, Heurteaux C, Lazdunski M. (1997) A proton-gated cation channel involved in acid-sensing. Nature.386: 173-177.
- Krishtal O. (2003) The ASICs: signaling molecules? Modulators? Trends Neurosci. 26(9): 477-483.
- Yermolaieva O, Leonard AS, Schnizler MK, Abboud FM, Welsh MJ (2004) Extracellular acidosis increases neuronal cell calcium by activating acid-sensing ion channel 1a. Proc. Natl Acad. Sci. USA. 101(17): 6752–7.
- Zha XM, Costa V, Harding AM, Reznikov L, Benson CJ, et al (2009) ASIC2 subunits target acid-sensing ion channels to the synapse via an association with PSD-95. J. Neurosci. 29(26): 8438–46.
- Rong C, Chen FH, Jiang S, Hu W, Wu FR, Chen TY, et al., (2012) Inhibition of acid-sensing ion channels by amiloride protects rat articular chondrocytes from acid-induced apoptosis via a mitochondrial-mediated pathway.Cell Biol Int. 36(7): 635-41.
- Chu XP, Xiong ZG. (2013) Acid-sensing ion channels in pathological conditions. Adv Exp Med Biol. 961: 419-31.
- Maria SC, Eleonora B, Paola T, Roberto P.Vasculitides (2015) The Complement System: a Comprehensive Review. Clinical Reviews in Allergy & Immunology. 49(3): 333-46.
- Fei WJ YL, Wu L, Deng F, Zhang Q, Hu B, et al., (2012) Expression of acidsensing ion channel in the vascular endothelial cells of Henoch–Schonlein Purpura patients. Acta Universitatis Medicinalis Anhui. 47: 189-193
- Ozen S, Pistorio A, Iusan SM, Bakkaloglu A, Herlin T, Brik R, et al., (2008) EULAR/PRINTO/PRES criteria for Henoch-Schonlein purpura, childhood polyarteritis nodosa, childhood Wegener granulomatosis and childhood Takayasu arteritis: Ankara . Part II: Final classification criteria. Ann Rheum Dis. 2010. 69: 798-806.
- Wang C, Liu X, Ye Z, Zhang J, Tang H, et al., (2009) Mesangial medium with IgA1 from IgA nephropathy inhibits nephrin expression in mouse podocytes. European Journal of Clinical Investigation. 39(7): 561-7.
- Zeng C, Ying T, Liu X, Peng H, Zhang H, et al., (2009) Serum IgA1 from patients with IgA nephropathy up-regulates integrin-linked kinase synthesis and inhibits adhesive capacity in podocytes through indirect pathways. Clin Invest Med. 32(1): E20-7.
- Yang YH, Huang YH, Lin YL, Wan LC. Chuang YH, et al. (2006) Circulating IgA from acute stage of childhood Henoch-Schönlein purpura can enhance endothelial interleukin (IL)-8 production through MEK/ERK signalling pathway. Clinical and Experimental Immunology. 144(2): 247-253
- Boehme MW, Galle P, Stremmel W (2002) Kinetics of thrombomodulin release and endothelial cell injury by neutrophil-derived proteases and oxygen radicals. Immunology. 107(3): 340-9.
- Sanchez FA, Rana R, Gonzalez FG, et al., (2011) Functional Significance of Cytosolic Endothelial Nitric-oxide Synthase (eNOS) regulation of hyperpermeability. J Biol Chem 2011. 286(35): 30409–30414.
- Bamburg JR (1999) Proteins of the ADF/cofilin family: essential regulators of actin dynamics. Annu Rev Cell Dev Biol. 15: 185-230.
- Carlier MF, Pantaloni D (1997) Control of actin dynamics in cell motility. J Mol Biol. 269(4): 459-67.
- Chen CH, Ho HH, Wu ML, Layne MD, Yet SF (2014) Modulation of cysteine-rich protein 2 expression in vascular injury and atherosclerosis. Mol Biol Rep. 41(11): 7033-41.
- Kee HJ, Kim GR, Cho SN, Kwon JS, Ahn Y, et al., (2014) miR-18a-5p MicroRNA Increases Vascular Smooth Muscle Cell Differentiation by Downregulating Syndecan4. Korean Circ J. 44(4): 255-63.
- Grifoni SC, Jernigan NL, Hamilton G, Drummond HA ( 2008) ASIC proteins regulate smooth muscle cell migration. Microvasc Res. 75(2): 202-10.





