SEB/OVA Facilitated TIM4 Expression in Intestinal Antigen Presenting Cells via Acetylation of Histone H3
Yang B1,2, Mo L1,3, Zeng L1, Li XX1, Qiu S3, Liu Z1, Yang P1*
1 The Research Center of Allergy and Immunology, Shenzhen University School of Medicine, Shenzhen, China.
2 Key Laboratory of Optoelectronic Devices and Systems of Ministry of Education and Guangdong Province, College of Optoelectronic Engineering,
Shenzhen University, Shenzhen, China.
3 Long gang ENT Hospital, Shenzhen, China.
*Corresponding Author
Pingchang Yang,
The Research Center of Allergy and Immunology,
Shenzhen University School of Medicine, Shenzhen 518060, China.
Tel: +86-755-86671907
Fax: +86-755-86671906
E-mail: pcy2356@163.com
Received: June 06, 2016; Accepted: June 23, 2016; Published: June 30, 2016
Citation: Yang B, et al., (2016) SEB/OVA Facilitated TIM4 Expression in Intestinal Antigen Presenting Cells via Acetylation of Histone H3. Int J Clin Med Allergy. 4(1), 38-43. doi: dx.doi.org/10.19070/2332-2799-150009
Copyright: Yang P© 2016. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
Background: Various animal models have been established to dictate the pathogenesis of food allergy and each one has its own limitation. This study aimed to investigate the detailed molecular mechanisms underlying Staphylococcal enterotoxin B (SEB)/ovalbumin (OVA)-induced allergic reaction in mouse intestine.
Methods: Using SEB/OVA-sensitized food allergic model, the effect of SEB/OVA on Th1/Th2 cytokine production was determined by ELISA. Mean while, the gene and protein expression of TIM4 in intestinal antigen presenting cells (APCs) was evaluated by PCR and Western blot, respectively. The inducible effect of SEB/OVA-primed APCs on TIM1 expression of CD4+ T cell was also investigated. Finally, the acetylation of histones induced by SEB/OVA was analyzed by Western blot.
Results: We demonstrated that SEB/OVA showed dual effect on Th1/Th2 responses. The mRNA and protein expression of TIM4 in intestinal APCs was significantly elevated by SEB/OVA sensitization, which could further induce TIM1 expression on CD4+ T cell. The enhancement of TIM4 was mediated by acetylation of histone H3.
Conclusion: The present study demonstrated that the effect of SEB/OVA on Th0 cell differentiation is dependent on its concentration. Furthermore, SEB/OVA induced TIM4 up-regulation may be via acetylation of histone H3.
2.Methods
2.1.Animals
2.2.Food allergic model
2.3.ELISA
2.4.Using chamber
2.3.Electron photomicrographs of mast cells
2.4.Immune cells isolation and purification
2.5.Real-time RT-PCR
2.6.Fluorescent in situ hybridization
2.7.Western blotting
2.8.Immuno fluorescence staining
2.9.Statistical analysis
3.Results
3.1.SEB/OVA showed dual effects on Th1/Th2 response in the intestine
3.2.Low dosages of SEB/OVA inducedfood allergic symptomsin mice
3.3.SEB/OVA facilitated TIM4 expression in mouse intestinal APCs
3.4.TIM1 expression in intestinal CD4+ T cells was up-regulated by SEB/OVA-primed APCs
3.5.SEB/OVA enhanced histone H3 acetylationat residue of lysine 9
4.Discussion
5.Acknowledgement
6.References
Introduction
Food allergy (FA) is a rapidly growing worldwide health concern, with the incidence as high as 4-6% in adults [1]. Yet the etiology of FA remains unclear. Skewed Th2 polarization and enhanced antigen specific-IgE are typical features of food allergy [2]. Currently, there is no remedy available to cure FA except avoiding eating antigens. Different animal models, with each one have its limitation, have been introduced to unravel the pathogenesis of FA [3]. Among them, SEB/OVA-sensitized murine model is a good choice, which recapitulates many clinical features of food allergy that are not seen in other models [4]. Thus, the underlying mechanism dictating SEB/OVA mediated-food allergy needs to be elucidated.
SEB is the most frequent contamination produced by the bacterium Staphylococcus aureus, which exist in food, such as milk/dairy products, eggs, meats, shellfish/fish, and the other food allergens. Chronic rhinosinusitis-derived SEB could be swallowed down to the gastrointestinal tract and degraded into small peptides, such as Ssp (2.5 kDa), which functioned as hapten to facilitate OVA uptake in dendritic cells (DCs) [5]. It has been shown that SEB is able to activate T cells and facilitate production of IL-4, -5, and -13 [6, 7]. In the presence of SEB, TIM4 was inducible in naïve B cells [8] or human DCs [9] in vitro, which could further potentiate naïve CD4+ T cell differentiate into Th2 cells. Liu et al.,[10] demonstrated that SEB might be transported into DCs via TLR2. Furthermore, in an experimental food allergic model, exposure to SEB/OVA increased TIM4 expression on DC, thereby promoting Th2 polarization [11, 12]. Moreover, SEB may be also involved in the differentiation of regulatory T cells (Treg) [13] and influences the DNA methylation pattern [14]. Tight junction protein claudin-2 (Cldn2), which could facilitate antigen to transport across epithelial barrier, was demonstrated to be stimulated by SEB [15]. However, the detailed molecular mechanism by which SEB regulates TIM4 expressionis not known.
In the present study, the diverse effect of SEB/OVA on T cell differentiation and intestinal hypersensitivity was determined in a mouse model. We found that SEB/OVA induced-up-regulation of TIM4 was related to acetylation of histone H3.
Methods
A 70-year-old female with polymyalgia rheumatica on corticosteroids for 2 years presented with pruritic rash. The rash began on her face and spread to her scalp, chest, upper and lower extremities. The patient self- increased her prednisone from 7mg to 40mg a day with no relief and experienced subjective fevers,chills,malaise, increased thirst and anorexia requiring hospitalization. Medications on admission included loraze pam, hydrochlorothiazade/ triamterene, fluticasone, tiotropium bromide, alendronate, ibuprofen, prednisone, and hydroxychloroquine. The patient was seen by Dermatology who noted diffuse erythematous pustular lesions (Figure I) on her scalp (Figure II), chest (Figure III), abdomen (Figure IV), and back (Figure V) and performed two skin biopsies. The differential diagnosis included Steven-Johnson’s syndrome (SJS), drug rash with eosinophilia and systemic symptoms (DRESS), acute generalized exanthematous pustulosis (AGEP), pustular psoriasis, and infectious etiologies.
All the animal experimental procedures were conducted according to the guidelines approved by the Experimental Animal Ethic Committee at Shenzhen University, and were carried out in accordance with the Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health (NIH publication no. 85-23, revised 1996). 6-8 weeks old female Bal b/c mice were housed in a SPF animal facility with a 12 h light-dark cycle and were free to access standard diet and water.
Balb/c mice were intragastrically introduced with 50 mg OVA and a designated dosage of SEB (Figure 1). After that, the mice were boosted with the same dosage of OVA/SEB orally on day 3, 6 and 10 after the first gavage. The mice were sacrificed on day 14 after the first gavage. Each group consisted of 6-10 mice.
Protein was extracted from the small intestine and cytokines of Th1 (IFN-γ) and Th2 (IL-4) was determined by commercial ELISA kits (R&D System Inc., MN, USA) according to the manufacturer’s instructions. Levels of TIM1 inculture media were determined by ELISA (anti-TIM1 antibody was purchased from R&D Systems), following the procedures published previously [16].
The intestinal epithelial permeability, which is an important indicator of intestinal barrier function, was examined with Using chamber as described [17]. Briefly, the jejunal segments were obtained from mice and incubated with Kreb’s solution in Using chambers. The chambers was bubbled with a mixture of 95% O2 and 5% CO2 and the temperature inside was maintained at 37°C by a circulating water bath. A pulse of 1mV was given to the tissues every five minutes. The change in Isc (indicated for tissue permeability) was monitored during the whole process.
Mast cell de granulation was observed with electronic microscopy as previously reported [16]. Briefly, jejunal segments from mouse intestine were excised and fixed with 2% glutar aldehyde and proprocessed routinely. Degranulation was defined as the absence of all or a portion of granular contents or reduced density of granular matrices. Ten mast cells were randomly selected from each mouse; 60 mast cells in total were analyzed for each group.
Mouse intestinal Peyer’s patches were collected from mice and single lymphocyte suspensions were prepared [18]. CD11b+/c+ cells and CD4+ CD25- T cells were purified from above cell suspensions using commercial reagent kits following the manufacturer’s instructions (Miltenyi Biotec, Germany). Briefly, the cells labeled with MACS CD11 c and CD 11b micro beads and passed through a VS+ column on a Super MACS magnetic separator. CD11b+/c+ cells were collected by removing the column from the magetic field and then flusing it with PBS containing 0.5% fetal calf serum.
For CD4+ T cell purification, the cell mixture was first labeled with MACS CD3 and CD4 micro beads for isolation of CD3+, CD4+ T cells. After that, the obtained cells were labeled with MACS CD25 micro beads and processed accordingly. The CD4+, CD25- T cells were collected and the purity of the cells was determined by flow cytometry. Only if the purity reached 98%, the cells were used for the following studies.
The total RNA was extracted from cells with TRIzol reagent (Invitrogen Corporation, CA, USA) and subjected to reverse transcription using a cDNA reverse transcription reagent kit (Qiagen, Mainz, Germany). The gene expressions of TIM4 or TIM1 were quantified by real-time qPCR using QuantiTect® SYBR® Green PCR Kit (Qiagen, Mainz, Germany). Fold changes in gene expression was calculated as 2ΔΔCT and the results were normalized to a percentage of the internal control β-actin. The primers of used in this study were as follows: TIM4, forward (5’-GCTGCTTCCAACAACAGTCA-3’) and reverse (5’-GTGATTGGATGCAGGCAGAG-3’); TIM1, forward (5’-ATGCCCATCTTCTGCTTG-3’) and reverse (5’-GTGCCATTCCAGTCTGTAGG-3’).
Fluorescent in situ hybridization of TIM4 was carried out as described else where [19]. Briefly, CD11b+/c+ cells were fixed by 4% paraformaldehyde routinely. After that, the cells were digested with proteinase K, and incubated with hybridization solution (containing 1 ng/μl fluoresce in conjugated TIM4 probe) at 37°C overnight. The cells were then immersed in 2X SSC (standard saline citrate) buffer at 45°C for twice and placed in 0.1XSSC for 10min. For signal observation, the cell were rinsed in PBS, air-dried, and visualized under a fluorescent microscope (Olympus, Japan).
After treatment, the total (for TIM4 or TIM1)or nuclear (for H3 acetylation) proteins were extracted from the CD11b+/c+ cells or CD4+ T cells. Equal amounts of protein (40μg/well) were separated by SDS-PAGE and transferred to PVDF membranes. The Membranes were incubated with primary antibodies (TIM4, TIM1, or H3 acetylation) at a specific concentration overnight at 4°C. The membranes were washed for three times and incubated with corresponding secondary antibodies at room temperature for 1 hour. The signals in membrane were then developed by ECL Plus Western Blotting Detection Reagents.
The expression of TIM1 in CD4+ T cells was visualized by immuno fluorescence staining routinely [20]. After treatment, the cells were fixed with cold acetone for 20 min, and blocked with 2% BSA solution for 60 minutes. The cells were incubated with the TIM1 antibody (1:200 dilution) for overnight at 4°C. After washing for three times with BSA solution, the cells were incubated with secondary antibody for 1 hour at room temperature. The nuclei were stained with propidium iodide for morphological viewing. The images were captured by Olympus microscope (Japan).
Data were expressed as the means ± S.D. Differences between groups were determined with Student’s t-test (2 groups)or ANOVA (3 or more groups). A P value less than 0.05 was considered to indicate significant differences.
Mice were treated with a serial dosage of SEB/OVA as shown in Figure 1, and proteins were extracted from the excised small intestinal tissues. The Th1 and Th2 functions were assessed by evaluation of the intestinal Th1 and Th2 cytokines with ELISA. As shown in Figure 2, co-administration of SEB and OVA increased intestinal IL-4 expression at low dosages of SEB(≤2.5 ug/mouse), while high dosages of SEB (2.5~40 ug/mouse) showed no effect on IL-4 expression. SEB/OVA significantly enhanced IFN-γ levels when SEB dosage reached and beyond 10 μg/mouse (Figure 2). The above results indicated that high dosages of SEB/OVA induced Th1 reactions whereas low dosages of SEB/OVA induced Th2 reactions in the intestine.


Figure 1. Frequency (%) of exposure to risk factors in control and asthma group during first life year.
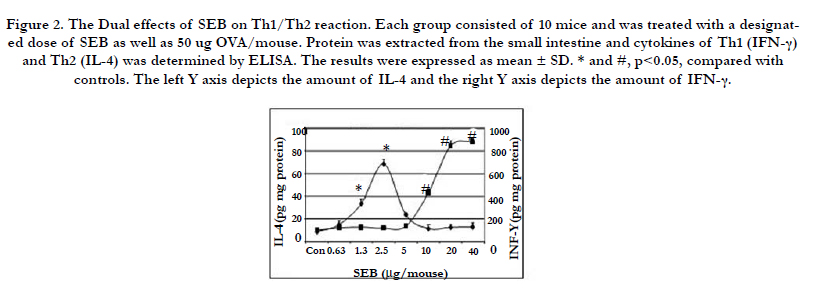
Figure 2. The Dual effects of SEB on Th1/Th2 reaction. Each group consisted of 10 mice and was treated with a designated dose of SEB as well as 50 ug OVA/mouse. Protein was extracted from the small intestine and cytokines of Th1 (IFN-γ) and Th2 (IL-4) was determined by ELISA. The results were expressed as mean ± SD. * and #, p<0.05, compared with controls. The left Y axis depicts the amount of IL-4 and the right Y axis depicts the amount of IFN-γ.
Next, we tested whether low dosages of SEB/OVA could induce food allergic symptoms in mice. As shown in Figure 3, the groups of mice treated with SEB/OVA (2.0~4.0 ug/mouse) had significantly more serum OVA-specific IgE (Figure 3A), higher Isc in response to the antigen challenge (OVA) in Using chambers (Figure 3B), and more extensive intestinal mucosal mast cell activation in a dose-dependent manner (Figure 3C-E). Neither did SEB nor OVA alone induce any signs of the intestinal hypersensitivity.
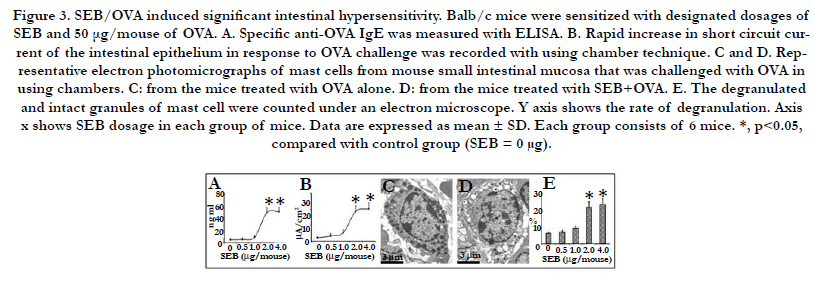
Figure 3. SEB/OVA induced significant intestinal hypersensitivity. Balb/c mice were sensitized with designated dosages of SEB and 50 μg/mouse of OVA. A. Specific anti-OVA IgE was measured with ELISA. B. Rapid increase in short circuit current of the intestinal epithelium in response to OVA challenge was recorded with using chamber technique. C and D. Representative electron photomicrographs of mast cells from mouse small intestinal mucosa that was challenged with OVA in using chambers. C: from the mice treated with OVA alone. D: from the mice treated with SEB+OVA. E. The degranulated and intact granules of mast cell were counted under an electron microscope. Y axis shows the rate of degranulation. Axis x shows SEB dosage in each group of mice. Data are expressed as mean ± SD. Each group consists of 6 mice. *, p<0.05, compared with control group (SEB = 0 μg).
The mature CD11b+/c+ APCs were treated with SEB and/or OVA. After 4-day incubation, the cells were harvested for evaluation of TIM4 expression level. TIM4 mRNA in the APCs was visualized in situ with confocal microscopy presenting in green color (Figure 4 A-D). Moreover, the mRNA expression level of TIM-4 was quantified by RT-PCR (Figure 4 E). The results showed thatSEB/OVA significantly increased mRNA level of TIM4 in APCs. Meanwhile, the protein level of TIM4 was significantly up-regu-lated by stimulation with SEB+OVA, but not with SEB alone or OVA alone (Figure 4F).
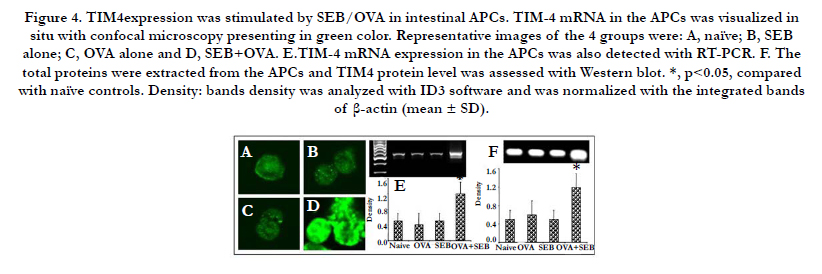
Figure 4. TIM4expression was stimulated by SEB/OVA in intestinal APCs. TIM-4 mRNA in the APCs was visualized in situ with confocal microscopy presenting in green color. Representative images of the 4 groups were: A, naïve; B, SEB alone; C, OVA alone and D, SEB+OVA. E.TIM-4 mRNA expression in the APCs was also detected with RT-PCR. F. The total proteins were extracted from the APCs and TIM4 protein level was assessed with Western blot. *, p<0.05, compared with naïve controls. Density: bands density was analyzed with ID3 software and was normalized with the integrated bands of β-actin (mean ± SD).
The interaction of TIM4 and TIM1 contributed to the development of intestinal Th2 polarization [16, 21, 22]. Thus, the direct inducible effect of SEB/OVA-primed APCs on TIM1 expression in intestinal CD4+ T cells was investigated in the present study. The immuno histochemistry images showed that TIM1 expression level was significantly higher in intestinal CD4+ T cells, which were cocultured with SEB/OVA-primed APCs (Figure 5 A and B). SEB/OVA-primed APCs also stimulated the secretion of TIM1 in CD4+ T cells (Figure 5C). Further analysis showed that both mRNA and protein expression levels of TIM1 in CD4+ T cells were increased after incubation with SEB/OVA-primed APCs(Figure 5 D and E).
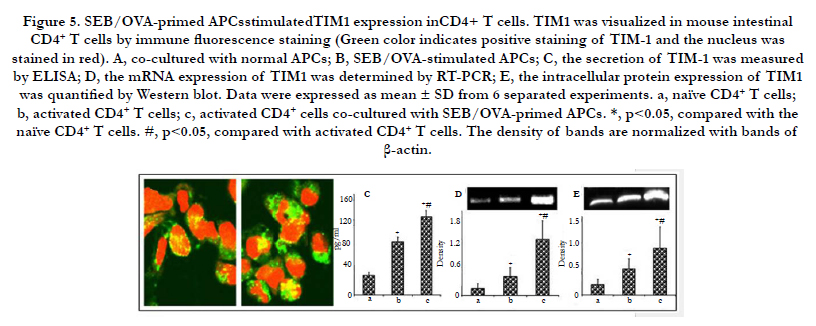
Figure 5. SEB/OVA-primed APCsstimulatedTIM1 expression inCD4+ T cells. TIM1 was visualized in mouse intestinal CD4+ T cells by immune fluorescence staining (Green color indicates positive staining of TIM-1 and the nucleus was stained in red). A, co-cultured with normal APCs; B, SEB/OVA-stimulated APCs; C, the secretion of TIM-1 was measured by ELISA; D, the mRNA expression of TIM1 was determined by RT-PCR; E, the intracellular protein expression of TIM1 was quantified by Western blot. Data were expressed as mean ± SD from 6 separated experiments. a, naïve CD4+ T cells; b, activated CD4+ T cells; c, activated CD4+ cells co-cultured with SEB/OVA-primed APCs. *, p<0.05, compared with the naïve CD4+ T cells. #, p<0.05, compared with activated CD4+ T cells. The density of bands are normalized with bands of β-actin.
Acetylation of the core histones at N-terminal correlates with enhanced transcription level, which is regulated by a balance between histone acetyl transferase and histone deacetylase. We have found that SEB/OVA increased TIM4 expression in the intestinal APCs, implying that SEB/OVA may be involved in the gene transcription of TIM4. Therefore, the nuclear proteins were extracted from CD11b+/c+ cells and subjected to Western blot to evaluate acetylated histone H3 with the polyclonal anti-acetylated histone H3 antibodies at specific lysine residues at 9, 14, 23 and 28 respectively. The results showed that SEB/OVA significantly increased the amount of acetylated histone H3 at lysine 9 in a dose-dependent manner (Figuer 6), but no changes at lysine 14 or 23 or 28 (data not shown). SEB or OVA alone showed no effect on histone acetylation. Histone acetylation plays a critical role in chromatin remodeling and gene expression, therefore, the increased levels of histone H3 acetylation at lysine9 can be considered as a mechanism by which SEB/OVA altered the gene expressions of TIM4.
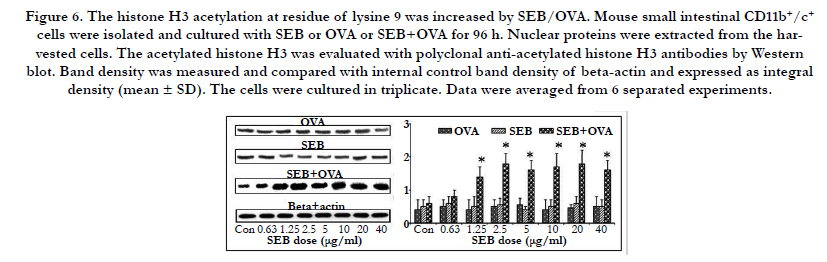
Figure 6. The histone H3 acetylation at residue of lysine 9 was increased by SEB/OVA. Mouse small intestinal CD11b+/c+ cells were isolated and cultured with SEB or OVA or SEB+OVA for 96 h. Nuclear proteins were extracted from the harvested cells. The acetylated histone H3 was evaluated with polyclonal anti-acetylated histone H3 antibodies by Western blot. Band density was measured and compared with internal control band density of beta-actin and expressed as integral density (mean ± SD). The cells were cultured in triplicate. Data were averaged from 6 separated experiments.
Discussion
The major findings in the present study include the followings:
(1) SEB showed dual effect on Th1/Th2 cytokines production in vivo;
(2) SEB/OVA-primed APC up-regulated TIM1 expressionin intestinal CD4+T cells; (3) SEB/OVA promoted TIM4 expression via acetylation of histone H3.
Growing evidences suggest a vital role of Staphylococcus aureus enterotoxins (SAE), consist of staphylococcal enterotoxins A, B, C, and D (SEA-D) and toxic shock syndrome toxin-1 (TSST- 1), in allergic diseases. For example, Staphylococcus aureus colonies were found to be strikingly higher (80-100%) in atopic eczema/ dermatitis syndrome patients (AEDS) than healthy controls (5-30%). SAE-specific IgE was detectable in the serum of 57% of AEDS patients [23]. Furthermore, SEB showed diverse effects on modulating functions of immune cells, including 1) entering DC via TLR2 [10, 24]; 2) modifying the properties of B cells [8]; 3) directly binding to the T cell receptors to activate T cells [16]; 4) inducing cytotoxic T cell differentiation [25]; 5) suppressing the expression of integrin αvβ6 in the nasal mucosa [26].
Aberrant Th2 polarization plays a critical role in the pathogenesis of allergic disorders. In order to uncover its unknown etiology, various animal models were introduced. Previous evidences from our and other laboratories all showed that SEB/OVA-sensitized food allergy model is a validated animal model to improve our understanding of its underlying mechanism and for establishing potential candidate for developing therapies [4]. This study provides the first evidence that SEB has dual effect on naïve CD4+ T cell differentiation. SEB promoted Th2 polarization only if its dose is lower than 2.5 ug/mouse, while high dose treatment showed opposite effect. This result is of great importance to instruct people who would like to apply this model in their study.
Recently, our group reported that DCs express TIM4 that ligates TIM1 on Th0 cells to promote Th2 cell development [12, 16]. We also demonstrated that SEB could promote Th2 production via up-regulation of TIM4 on cell surface of DC [9], while the underlying molecular mechanism remains unknown. The present study further revealed that only low doses of SEB (<2.5 ug/mouse) stimulated Th2 reaction and SEB-facilitated TIM4 enhancement was possibly via acetylation of histone H3.
Acknowledgement
This study was supported by grants from the innovation of science and Technology Commission of Shenzhen Municipality (JCYJ20140418095735611 and ZDSYS201506050935272), the Natural Science Foundation of China (81373176, 31570932, 31400856, 81300292,81571790 and 81501573).
References
- Bischoff SC (2006) Food allergies. Curr Gastroenterol Rep. 8(5): 374-82.
- Tournoy KG, Provoost S, Van Hove C, Joos G (2006) The role of immune tolerance in asthma pathogenesis. Curr Allergy Asthma Rep, 6(5): 437-43.
- Abril-Gil M, Garcia-Just A, Pérez-Cano FJ, Franch À, Castell M (2015) Development and characterization of an effective food allergy model in Brown Norway rats. PLoS One. 10(4): e0125314.
- Ganeshan K, Neilsen CV, Hadsaitong A, Schleimer RP, Luo X, et al., (2009) Impairing oral tolerance promotes allergy and anaphylaxis: a new murine food allergy model. J Allergy Clin Immunol. 123(1):231-238 e4.
- Yang SB, Li TL, Chen X, An YF, Zhao CQ et al., (2013) Staphylococcal enterotoxin B-derived haptens promote sensitization. Cell Mol Immunol. 10(1): 78-83.
- Patou J, Gevaert P, Van Zele T, Holtappels G, van Cauwenberge P, et al., (2008) Staphylococcus aureus enterotoxin B, protein A, and lipoteichoic acid stimulations in nasal polyps. J Allergy Clin Immunol. 121(1): 110-5.
- Kim DW, Khalmuratova R, Hur DG, Jeon SY, Kim SW, et al., (2011) Staphylococcus aureus enterotoxin B contributes to induction of nasal polypoid lesions in an allergic rhinosinusitis murine model. Am J Rhinol Allergy.25(6): e255-61.
- Qiu S, Du Y, Duan X, Geng X, Xie J, et al., (2012) B cell immunity in allergic nasal mucosa induces T helper 2 cell differentiation. J Clin Immunol.32(4): 886-95.
- Liu T, He SH, Zheng PY, Zhang TY, Wang BQ, et al., (2007) Staphylococcal enterotoxin B increases TIM4 expression in human dendritic cells that drives naive CD4 T cells to differentiate into Th2 cells. Mol Immunol,44(14): 3580-7.
- Huvenne W, Callebaut I, Plantinga M, Vanoirbeek JA, Krysko O, et al., (2010) Staphylococcus aureus enterotoxin B facilitates allergic sensitization in experimental asthma. Clin Exp Allergy, 40(7): 1079-90.
- Tao Liu, Bin-Quan Wang, Peng-Yuan Zheng, Shao-Heng He, Ping-Chang Yang, (2006) Rhinosinusitis derived Staphylococcal enterotoxin B plays a possible role in pathogenesis of food allergy. BMC Gastroenterol, 6:24.
- Yang PC1, Xing Z, Berin CM, Soderholm JD, Feng BS, et al., (2007) TIM- 4 expressed by mucosal dendritic cells plays a critical role in food antigenspecific Th2 differentiation and intestinal allergy. Gastroenterology, 133(5):1522-33.
- Cho SN1, Song CH, Jin J, Kim SH, Rha KS ., et al., (2014) Role of staphylococcal enterotoxin B on the differentiation of regulatory T cells in nasal polyposis. Am J Rhinol Allergy. 28(1): 17-24.
- Pérez-Novo CA, Zhang Y, Denil S, Trooskens G, De Meyer T, et al., (2013) Staphylococcal enterotoxin B influences the DNA methylation pattern in nasal polyp tissue: a preliminary study. Allergy Asthma Clin Immunol. 9(1): 48.
- Liu X, Yang G, Geng XR, Cao Y, Li N,, et al., (2013) Microbial products induce claudin-2 to compromise gut epithelial barrier function. PLoS One. 8(8): e68547.
- Feng BS, Chen X, He SH, Zheng PY, Foster J, et al., (2008) Disruption of T-cell immunoglobulin and mucin domain molecule (TIM)-1/TIM4 interaction as a therapeutic strategy in a dendritic cell-induced peanut allergy model. J Allergy Clin Immunol. 122(1): 55-61.
- Ping-Chang Yang, Ya-Hong Tu, Mary H. Perdue, Christine Oluwole, Stevie Struiksma (2009) Regulatory effect of heat shock protein 70 in stressinduced rat intestinal epithelial barrier dysfunction. N Am J Med Sci, 1(1):9-15.
- Lyscom, N, M.J. Brueton (1982) Intraepithelial, lamina propria and Peyer's patch lymphocytes of the rat small intestine: isolation and characterization in terms of immunoglobulin markers and receptors for monoclonal antibodies.Immunology. 45(4):. 775-83.
- Karteris E, Randeva HS, Grammatopoulos DK, Jaffe RB, Hillhouse EW (2001) Expression and coupling characteristics of the CRH and orexin type 2 receptors in human fetal adrenals. J Clin Endocrinol Metab. 86(9): 4512-9.
- Collins-Hooper H, Sartori R, Macharia R, Visanuvimol K, Foster K , et al., (2014) Propeptide-mediated inhibition of myostatin increases muscle mass through inhibiting proteolytic pathways in aged mice. J Gerontol A Biol Sci Med Sci. 69(9): 1049-59.
- Kim HY, Chang YJ, Chuang YT, Lee HH, Kasahara DI, et al., (2013) T-cell immunoglobulin and mucin domain 1 deficiency eliminates airway hyperreactivity triggered by the recognition of airway cell death. J Allergy Clin Immunol. 132(2):. 414-25.
- Zhao CQ, Li TL, He SH, Chen X, An YF, et al.,(2010) Specific immunotherapy suppresses Th2 responses via modulating TIM1/TIM4 interaction on dendritic cells. Allergy. 65(8): 986-95.
- Na SY, Roh JY, Kim JM, Tamang MD, Lee JR. (2012) Analysis of Colonization and Genotyping of the Exotoxins of Staphylococcus aureus in Patients with Atopic Dermatitis. Ann Dermatol. 24(4): 413-9.
- Liu T, Xie C, Chen X, Zhao F, Liu AM, et al.,(2010) Role of muscarinic receptor activation in regulating immune cell activity in nasal mucosa. Allergy. 65(8): 969-77.
- Qiu S, Du Y, Duan X, Geng X, Xie J, et al., (2011) Cytotoxic T lymphocytes mediate chronic inflammation of the nasal mucosa of patients with atypical allergic rhinitis. N Am J Med Sci. 3(8): 378-83.
- Liu T, Liang X, Li TL, Ma J, Yang JF, et al.,(2012) Staphylococcal enterotoxin B compromises the immune tolerant status in the airway mucosa. Clin Exp Allergy. 42(3): 375-82.





