Methods of Studying the Social Structure of Early Hominins: A Mini Review
Al-Imam A*
Department of Anatomy and Cellular Biology, College of Medicine University of Baghdad, Iraq.
*Corresponding Author
Dr. Ahmed Al-Imam
Department of Anatomy and Cellular Biology,
College of Medicine University of Baghdad, Iraq.
E-mail: tesla1452@gmail.com
Received: July 24, 2017; Accepted: August 29, 2017; Published: August 31, 2017
Citation: Al-Imam A. Methods of Studying the Social Structure of Early Hominins: A Mini Review. Int J Anat Appl Physiol. 2017;3(2):69-73. doi: dx.doi.org/10.19070/2572-7451-1700011
Copyright:Al-Imam A© 2017. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
There has been numerous attempts by anthropologists and paleoanthropologist to study the social structure of early hominins, many of these were based on three main modalities of research; non-human primate models, behavioural ecology, and phylogenetic models. Non-human primate models are considered as the archetypal analytic approach of social structure. On the other hand, behavioural ecology emphasises on the shared behavioural patterns in between our specie, the early hominin species, and the species of African apes. The 3rd approach, the phylogenetic modelling, is considered to be the most novel method; It relies on the analysis of phylogenetic constraints in a particular ecosystem; which seems to enforce a specific type of social structure; it represents a responsive interaction between the; environment, evolutionary biology, and ultimately the social fabric.
2.Background
3.Nonhuman Primate Models
4.Historical Nonhuman Primate Studies
5.Phylogenetic Models
6.Behavioral Ecology
7.Conclusion
8.References
KeyWords
Physical Anthropology; Bipedalism; Locomotion; Hominidae; Hominin; Genus Homo; Australopithecus; Critical Analysis; Anatomy; Comparative Anatomy; Pelvis; Femur; Gluteal musculature.
Background
Hominidae includes the great apes; chimpanzees, bonobos, orangutans, gorillas, and humans. Formerly, humans, both extinct and extant, were placed in Hominidae, while great apes were put in a different family, Pongidae family [1, 2]. Despite the presence of landmark physiological differences in between humans and great apes, for example, the relative longer reproductive life span in chimpanzees females, both morphological and molecular studies confirmed that humans and chimpanzees are closely related and interlinked by a common ancestor at approximately six million ears ago (Mya) [1-3].
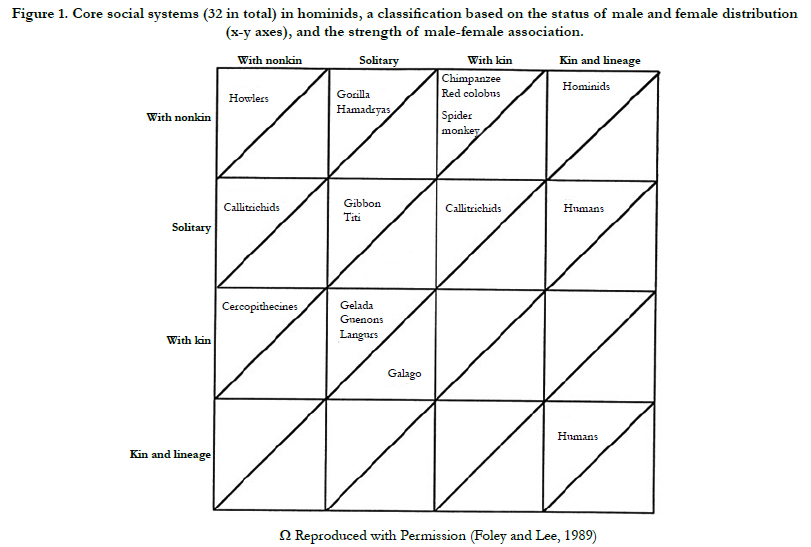
Nevertheless, it is agreed that some traits are unique for human, these are; imitation, language and verbal skills, obligate bipedal locomotion, stable supine posture, and imagination [4-6]. The social structure of early hominin cannot be easily inferred as in the case of physical anthropology and archaeology. Hence, anthropologists use three main methods to infer data in relation to the social fabric of early hominins. Those methods include; nonhuman primate models, phylogenetic history, and behavioural ecology principles [7-10]. The landmark paper by Foley and Lee [11], discussed the presence of 32 core social systems in hominids (Figure 1); the classification is based on the status of male and female distribution, and the strength of male-female association [11].
Nonhuman Primate Models
Modelling based on non-human primates is regarded as the archetype approach for social-marital structure studies. The social structure in apes can be diverse, for example; monogamy (Gibbons), uni-male polygyny (Orangutan), uni-male polygyny (gorilla), and multi-male polygyny (chimpanzee). Although the savannah baboon monkeys are distantly related to hominins, they do have a profoundly similar social structure to early hominins due to the similar nature of their environments, which is represented by the open terrains of the Savannah [1]. The chimpanzee was also proposed as a model for studying the social structure in early hominins; chimpanzees share a common ancestry with early hominins; chimps may also use tools and hunt occasionally. However, chimpanzees are extant (living) models, and they differ anatomically from early hominins, for example, the different facial and dental anatomy when compared to the Australopithecines (afarensis, robustus, and africanus). Hence, the extinct hominins behavioural and social patterns would also be different from those of the chimpanzees [1, 12, 13]. In 1978, the pygmy chimpanzee known as bonobos (Pan paniscus) became an adopted model; it shares similar overall body proportions with an early hominin species known as Australopithecus afarensis (Lucy’s specie); this hominin existed at around 3.2 Mya in east Africa. Furthermore, the bonobo's society is an egalitarian society promoting equality among its members, though it is more female-centered in which sex is a substitute for aggression and violence. Accordingly, Pan paniscus is considered as amore acceptable model (than Pan troglodytes) [1, 14].
Historical Nonhuman Primate Studies
Chimpanzees (common chimpanzee) and bonobos (pygmy chimpanzee) were used as non-human primate models to understand the behaviour and social structure of early hominins. Primatologists who pioneered the fields experiments include Tetsuro Matsuzawa, Jane Godall, and Dian Fossey. Each scientist has done intensive work on non-human primates to deduct the archaic behaviour of early hominins [1, 15, 16].
According to Tetsuro Matsuzawa, Hominidae members are considered to have a protoculture. Culture and protoculture require three basic elements; emergence, transmission, and modification [17]. Protoculture emergence has been fictionally depicted in Rise of the Planet of the Apes in which a leader chimpanzee named Caesar begins a revolution against humans [18]. Several examples of intriguing protoculture have been observed in chimpanzees (Pan troglodytes) including; Sweet potato washing in snow (Japanese) monkeys in Koshima (Japan), hot-bathing snow monkeys in shiga heights (Japan), and the extra-large cluster of Shodoshima snow monkeys in Saru Dango (Japan). Another interesting example is the deer-riding behaviour noticed in exclusively in a snow monkey sub-species (Macaca fuscata yakui) in Yakushima (Japan); the emergence of this commensalism-like behaviour (deer-riding) was first observed in a monkey nicknamed Rodeo [16, 19, 20]. Behavioural patterns have also been approached via the master-apprenticeship method, as studied by Matsuzawa with Ai and Ayumu (chimpanzees’ nicknames) and by Tomonaga with Chloe and Cleo [16, 17].
Behavioural studies can be either observational or experimental, in the laboratory or the natural habitat of a primate; these have been extensively studied by eminent primatologist including Jane Godall and Dian Fosse, while the systematic field experiments are deemed to be pioneered by Tetsuro Matsuzawa [16]. Matsuzawa and colleagues have done numerous and remarkable behavioural, cognitive, and social studies in common chimpanzee, including the Ai project (1978), other studies in Skylab (2008) and Kumamoto Sanctuary (2011), field studies in Bossou-Africa, and in other regions of Central and West Africa [21-26]. Most of the studies were conducted either in the east or central Africa, while fewer ones were carried out in West Africa.
Phylogenetic Models
Phylogenetic models are based on common behavioural patterns in between the species of the African apes and humans. Wrangham and colleagues examined different behavioural traits (fourteen traits) including male-female interactions, social structure, and intergroup aggression. It was deducted that If a trait exists in all four species, it is likely to exist in the shared (common) ancestor [27, 28]. On the other hand, If the four species differ regarding specific trait, then nothing can be inferred about the common ancestor [27-29]. Wrangham also concluded that the common ancestor of African apes and hominins had closed social networks characterised by male-centered dominion, polygynous males, female exogamy, and no inter-females alliances. However, Lovejoy (1981) proposed that Australopithecus afarensis were non-hostile and monogamous [1].
Behavioral Ecology
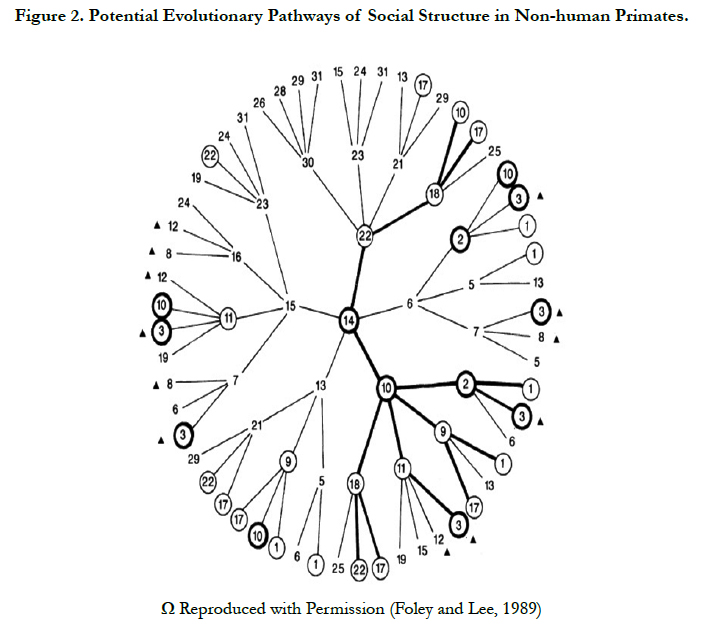
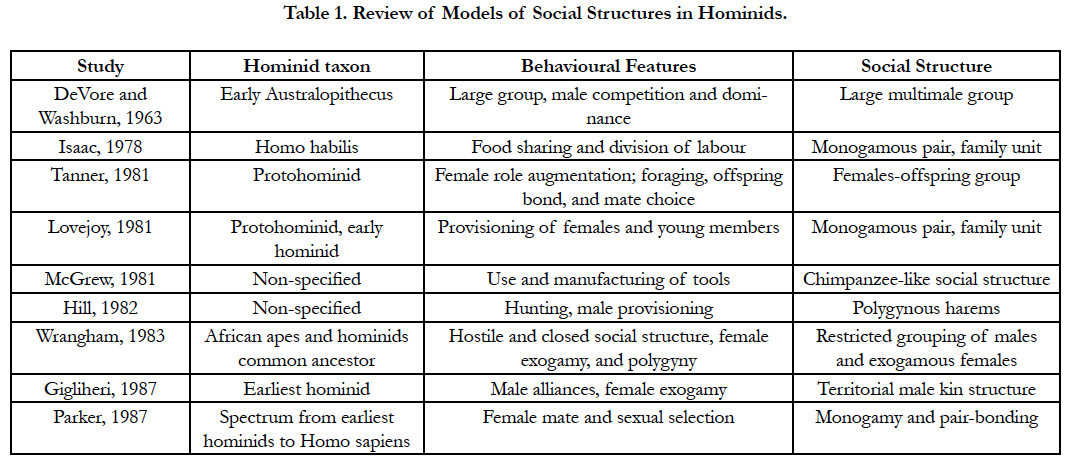
The third and the newest approach is based on the behavioural ecology principles (Table 1); it determines how the social structure can be modified in correspondence with the existing environment. It is an analysis of phylogenetic constraints in a particular ecosystem; it further implies that both ancestral anatomy and the social structure will restrain the potential social evolutionary pathway. In other words, there are limited and predicted pathways (Figure 2) for an ecosystem-driven modification in social structure [11, 30]. The social structure is highly variable among apes (Figure 1) including; solitary individuals (orangutans), monogamous pairs and family units (Gibbons), single male units with small numbers of non-kin females (gorillas), complex fission–fusion societies (chimpanzees), and some degree of male kin bonding (most African apes) [1, [11, 30].

Approximately 10 Mya (end of Miocene) the steadily cooling climate was responsible for reducing the dense forests, which eventually led to a drier habitat with sparse food distribution in East Africa, particularly in the region of the great rift valley [1, 9]. This ecological modification favoured the evolution of a chimplike social structure of dispersed females with their offspring and kin-related males for defensive-offensive warfare against other competing groups. The earliest Hominin social fabric occurred in parallel with enhanced male kinship alliances within the geography of an open terrain (the Savannah). The persistent cooling trend promoted these open terrains and increased predatory threats from other species. As a result, the social structure demanded an increment in group size (more members) to enhance survival; those large groups required strong males alliances with other related males (based on kinship). Hence, early hominins are most likely to live in big groups of men and women, up to 200 individuals altogether with their offsprings, with the males being closely related to each other. The females would be given the task to forage for food resources in the open terrains, where they would enhance their defence from predators by associating with other females, and one or more strong males within their group [1, 9-11].
Approximately 3 to 1 Mya, hominin species developed a degree of anatomic diversities as a form of adaptive subsistence, for instance, Australopithecus robustus fed on hard plant-based low-quality diet found in the dispersed patches of the savannah. Bonds between males became weaker due to more intense competition for food resources; this was also accompanied by sexual dimorphism (gender-based body size discrepancies). On the other hand, the later Homo erectus evolved an augmented (endo)- cranial capacity and a reduced dental apparatus. Moreover, the erectines who also lived in a similar tropical environment and the open savannah, adopted an entirely different subsistence strategy from the robusts, they increased the caloric meat consumption to feed their hungry brains, this was promoted and accompanied by the emergence of tool-making abilities which are basically a manifestation of an advanced brain power and a superior cognitive skills. However, they also had to roam for meaty food resources to considerable distances, which led to the compromise of male ability to defend the females. As a result, those males had to rely on solid alliances by strengthening their kinship bonding for an effective territorial defence against external threats [27-30].
Conclusion
The social structure for early hominins is complex, and cannot be easily inferred; The complexity of the social fabric of early hominins is paralleled only by the complex nature of its analytic models and tools. Several models exist; each of these is based on a specific analytic method or a combination of methods including; nonhuman primate models, behavioural ecology, and phylogenetic models. Several attempts were carried out by anthropologist and paleoanthropologist in this field. However, these models are neitherfully accurate nor foolproof. Hence, an integrative approach to all three models might be mandatory. Behavioural ecology appears to be special; its uniqueness arises from the Integrative observation and analysis of the ecosystem, the anatomic and evolutionary adaptations, and the available possibilities of social structure. Behavioural ecology is responsive to the interaction of the environment versus the evolutionary adaptations and social organisation of the early hominins.
References
- Lewin R. Human evolution: an illustrated introduction. 5th ed. New jersy:John Wiley & Sons; 2009 Feb 18.
- Smith BH, Tompkins RL. Toward a life history of the Hominidae. Ann Rev Anthropol. 1995 Oct;24(1):257-79.
- Hill A, Ward S. Origin of the Hominidae: the record of African large hominoid evolution between 14 My and 4 My. Am J Phys Anthropol. 1988 Jan 1;31(S9):49-83.
- Matsuzawa T. Comparative cognitive development. Dev sci. 2007 Jan 1;10(1):97-103.
- Saito A, Hayashi M, Takeshita H, Matsuzawa T. The Origin of Representational Drawing: A Comparison of Human Children and Chimpanzees. Child dev. 2014 Nov 1;85(6):2232-46.
- Sakuraba Y, Tomonaga M, Hayashi M. A new method of walking rehabilitation using cognitive tasks in an adult chimpanzee (Pan troglodytes) with a disability: a case study. Primates. 2016 Jul 1;57(3):403-12.
- Bogin B. The evolution of human childhood. Biosci. 1990 Jan 1;40(1):16-25.
- Crockett CM, Eisenberg JF, Smuts BB, Cheney DL, Seyfarth RM, Wrangham RW, et al. Primate societies. University of chicago press; 1987.
- Foley R. Humans before humanity: an evolutionary perspective. Blackwell; 1997.
- Kent George Cantine, Carr Robert. Comparative anatomy of the vertebrates. 9th ed. Newyork:McGraw-Hill; 2001.
- Foley RA, Lee PC. Finite social space, evolutionary pathways, and reconstructing hominid behavior. Sci. 1989 Feb 17;243(4893):901-906.
- Di Fiore A, Rendall D. Evolution of social organization: a reappraisal for primates by using phylogenetic methods. Proc Natl Acad Sci USA. 1994 Oct 11;91(21):9941-5.
- Lee PC. Social structure and evolution. In: Slater PJB, Halliday TR, editors. Behavior and evolution. Cambridge, UK: Cambridge University Press, 1994 Sep 15. 266–303.
- Zihlman AL, Cronin JE, Cramer DL, Sarich VM. Pygmy chimpanzee as a possible prototype for the common ancestor of humans, chimpanzees and gorillas. Nature. 1978 Oct 26;275(5682):744-6.
- Matsuzawa T (Kyoto University). Origins of the Human Mind. 2016 Feb 12. Available from https://www.edx.org/course/origins-human-mind-kyotoux-007x
- CICASP. Primate Research Institute, Kyoto University. The 50th Anniversary Symposium of Primate Research Institute of Kyoto University - Past, present and future of primatology. January 30th-31st,2017. PWS- Leading graduate programme in primatology and wildlife science. 2017 Feb 19.
- McGrew WC. The cultured chimpanzee: nonsense or breakthrough? Hum Ethol Bull. 2015 Jul 4;30(1):40-51.
- IMDb. Rise of the Planet of the Apes. 2011 Feb 20.
- Hirata S, Watanabe K, Masao K. “Sweet-potato washing” revisited. In: Primate origins of human cognition and behavior. Japan: Springer; 2008. 487-508.
- Kawai M. Newly-acquired pre-cultural behavior of the natural troop of Japanese monkeys on Koshima Islet. Primates. 1965 Aug 1;6(1):1-30.
- Biro D, Matsuzawa T. Chimpanzee numerical competence: cardinal and ordinal skills. In: Matsuzawa T, editor. Primate origins of human cognition and behavior. Japan: Springer; 2008. 199-225.
- Matsuzawa T. Evolution of the brain and social behavior in chimpanzees. Curr Opin Neurobiol. 2013 Jun 30;23(3):443-9.
- Matsuzawa T. Sociocognitive development in chimpanzees: a synthesis of laboratory work and fieldwork. In: Matsuzawa, Tomonaga M, Tanaka M, editors. Cognitive development in chimpanzees. Tokyo: Springer ; 2006.3-33.
- Matsuzawa T. Symbolic representation of number in chimpanzees. Curr opin neurobiol. 2009 Feb 28;19(1):92-8.
- Thompson ME, Jones JH, Pusey AE, Brewer-Marsden S, Goodall J, Marsden D, et al. Aging and fertility patterns in wild chimpanzees provide insights into the evolution of menopause. Curr Biol. 2007 Dec 18;17(24):2150-6.
- Tomonaga M, Matsuzawa T. Enumeration of briefly presented items by the chimpanzee (Pan troglodytes) and humans (Homo sapiens). Anim Learn Behav. 2002 May 1;30(2):143-57.
- Rodseth L, Wrangham RW, Harrigan AM, Smuts BB, Dare R, Fox R, et al. The human community as a primate society [and comments]. Curr Anthropol. 1991 Jun 1;32(3):221-54.
- Wrangham RW. Evolution of social structure. In: Smuts BB, Dorothy L, Seyfarth RM, Richard WR, editors. Primate societies. Chicago: The University of Chicago Press; 1987.
- Potts R. Reconstructions of early hominid socioecology: a critique of primate models. In: WG Kinzey, editor. The evolution of human behavior: Primate models. New york: Sunny press; 1987. 28-47.
- Foley RA. An evolutionary and chronological framework for human social behaviour. Proc Brit acad. 1996 Jan 1;88:95-118.





