Inhibitory - Proliferative Adaptation
Robert Skopec*
Researcher Analyst, AXON, Dubnik, Slovakia.
*Corresponding Author
Robert Skopec,
Researcher Analyst, AXON, Dubnik, Slovakia.
E-mail: zxcbnvm7@gmail.com
Received: July 21, 2016; Accepted: July 25, 2016; Published: August 04, 2016
Citation: Robert Skopec (2016) Inhibitory - Proliferative Adaptation. Int J Vaccines Res. 3(3), 19-22.
Copyright: Robert Skopec© 2016. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Abstract
We try to interpret origin and complexity of life in terms of ‘entropy dissipation’, and how the complexity of life could arise by ‘internal selection’ without the need of ‘Darwinian theory of natural selection’. Have physicists found the underlying science driving the origin and evolution of life ? It follows from the fundamental laws of physics. From this standpoint, there is one essential difference between living subjects and their corresponding inanimate carbon atoms: The former tend to be much better at capturing energy from their environment and dissipating that energy as heat. The mathematical formula, based on established physics, indicates that when a group of atoms is driven by an external source of energy and surrounded by a heat bath, will often gradually restructure itself in order to dissipate increasingly more energy. Under similar conditions, matter inexorably acquires the key physical attribute associated with life. From the perspective of the Prigogine- England physics, Darwinian evolution is only a special case of more general phenomenon.
2.The role of brain regions in modulating social stressors
3.Hierarchical arrangement of adaptation with brain at the top
4.The inhibitory-proliferative adaptation
5.Conclusions
6.References
Introduction
Theoretical results of physicists from Prigogine to England are generally considered valid. Their formula represents the driving force behind a class of phenomena in nature that includes life. A system of particles is driven by an oscillating force. Over time (top-down), the force triggers the formation of more bonds among the particles. At the heart of Prigogine-England theory is the second law of thermodynamics, the law of increasing entropy (the arrow of time) [3, 7]. The energy tends to disperse or spread out as time progresses. Entropy is the measure of this tendency. It increases as a matter of probability: There are more ways for energy to be spread out than for it to be concentrated. The entropy must increase over time in an isolated or “closed” system-an “open” system can keep its entropy low-that is, divide energy unevenly among atoms-by greatly increasing the entropy of its surroundings. In his influential 1944 monograph “What Is Life ?” physicist E. Schrὅdinger argued that this is what living systems must do. The overall entropy of the universe increases during photosynthesis as the sunlight dissipates, even as the plant prevents itself from decaying by maintaining an orderly internal structure. In the late 1990s, it was shown that entropy produced by a thermodynamic process corresponds to a simple ratio: the probability that the atoms will undergo that process divided by their probability of undergoing the reverse process. As entropy production increases, so does this ratio: A system’s behavior becomes more and more “irreversible”. Particles tend to dissipate more energy when they resonate with a driving force. “This means clumps of atoms surrounded by a bath at some temperature, like the atmosphere or the ocean, should tend over time to arrange themselves to resonate better and better with the sources of mechanical, electromagnetic or chemical work in their environments” [3].
The role of brain regions in modulating social stressors
The brain plays critical role in appraising social stressors, as well as in modulating the immune system’s response to stressors that involve social or physical threat. Differences of inflammatory responses to social stress depend on individual differences in activity of neural regions that process social threat-related information.
Stressors of social rejection up-regulate inflammatory activity. The neural regions involved in processing rejection-related distress relate to individuals magnitude of inflammatory responses to social stress.
General stress-response-controlled switch to error-prone DNA break (double-strand break, DSB) repair-dependent stress induced mutation, driven by spontaneous DNA breaks, is pathway that cells use and a major source of spontaneous mutation. Mechanisms that couple mutagenesis to stress responses allow cells to evolve rapidly and responsively to their environment. Furthermore, stress is not required, activation of the stress response is sufficient [10].
Self-replication (or reproduction-in biological terms), a necessary essential component for the evolution of life, is one such mechanism by which a system might dissipate an increasing amount of energy over time [16, 17]. The theoretical minimum amount of dissipation that can occur during the self-replication of RNA molecules and bacterial cells is very close to the actual amounts these systems dissipate when replicating.
According to Prigogine-England’s theory, the underlying principle driving whole process is dissipation-driven adaptation of matter [3,7]. This principle would apply to inanimate matter as well. Besides self-replication, greater structural organization is another means by which strongly driven systems ramp up their ability to dissipate energy. The distinction between living and nonliving matter is not sharp (see abiogenesis and the classic Miller experiment) [18]. This new facts will have impact on ecology, robotics and environment at all, including cancer microenvironment too. England argues that under certain conditions matter will spontaneously self-organize [3]. Research of the above process will include the biological and mathematical steps of this self-organization.
England’s theory could be tested by comparing cells with different mutations and looking for a correlation between the amount of energy the cells dissipate and their replication rates. Connection between England’s theory with Brenner’s microsphere constructions may show whether the theory correctly predicts which self-replication and self-assembly processes can occur, is probably a fundamental question in science today [3, 19].
Conventional wisdom holds that complex structures evolve from simpler ones, by step-by-step method, through a gradual evolutionary process, with Darwinian selection favoring intermediate forms along the way. But recently a growing number of scholars have proposed that complexity can arise also by other means-as a side effect, for example-even without natural selection to promote it. Studies suggest that random mutations that individually have no effect on an organism can fuel the emergence of complexity in a process named as constructive neutral evolution [20, 21].
Complexity can arise through other routes, because life has a built-in tendency to become more complex over time. It has been newly shown that random mutations arise, complexity emerges as a side effect, even without natural selection. For example, D. W. McShea and R. N. Brandon in their book Biology’s First Law [9] argued that bunch of parts that start out more or less the same should differentiate over time. When organisms reproduce, one or more of their genes may mutate. These mutations give rise to more type of the parts. If an organism has more parts, those units have an opportunity to become different. After a gene is accidentally copied, the duplicate may pick up mutations that the original does not share. If you start with a set of identical parts, they will tend to become increasingly different from one another. It means, the organism’s complexity will increase.
As complexity arises, it may help an organism survive better or have more offspring. If so, it will be favored to develop and spread through the population. The new copies will mutate. Natural selection will keep these mutations from spreading through populations. That is, organisms born with those traits will tend to die before reproducing, thus taking these traits out of circulation. Complexity arises mostly due to natural selection [16, 17].
Unlike standard evolutionary theory, we can see complexity increasing even in the absence of natural selection. It maintain a fundamental law of biology-perhaps its only one-the zero-force evolutionary law [9].
Consequently, natural selection is strong among the organisms living in wilder, eliminating mutations that make, for example, flies unable to cope with their many challenges. In sheltered environment of the labs, in contrast, natural selection is feeble. Lab-raised fruit flies are more complex than wild ones because their sheltered environment allows even disadvantageous mutations to spread. The zero-force evolutionary law makes a clear prediction: over their multiple use in research, the lab flies should have been less subject to the elimination of redundant (disadvantageous) mutations and thus should have become more complex then the wild ones [9].
An organism can exist without external selection-without the environment determining who wins and loses in the evolutionary race-but it can be subject to internal selection, which takes place within organisms. Freed from natural selection, organisms have reveled in complexity. For this type of evolution is an exact label “constructive-neutral evolution.” Experiments may decide why don’t we just have genes with the correct original sequence, making RNA editing unnecessary, etc. Further research will coax multiple biological theories to think beyond natural selection and to see the possibility that random mutation can fuel the evolution complexity on its own. Our point is that we don’t dismiss adaptation, we just don’t think it explains everything.
Hierarchical arrangement of adaptation with brain at the top
In hierarchical models causes in one level generate subordinate causes in lower level (sensory data per se are generated at the lowest level). Using the free energy effectively optimizes empirical priors, i.e. the probability of causes at one level is given those in the level above. Because empirical priors are linked hierarchically, they are informed by sensory data, enabling the brain to optimize its prior expectations online.
The free-energy construct was introduced into statistical physics to convert difficult probability-density integration problems into easier optimization problems. In the present context, free energy provides the answer to a fundamental question: how do self-organizing adaptive systems avoid surprising states ? [8] Our answer is: they can do this by optimizing (not only minimizing, suppressing, i.e. Darwinian selection) their free energy.
Physiological/biological complexity is the difference between the recognition density and the prior density on causes. The difference between the prior density (encodes beliefs about the state of the world before sensory data that are assimilated) and the recognition density (encodes posterior beliefs) [8]. The agent will selectively sample the sensory inputs that it expects: we anticipate what might be next and then try to confirm those expectations. So, when we are adapting to the environment, we conform to expectations [22, 23].
The units in the level above encode conditional expectations which optimize top-down predictions to explain away (to reduce) prediction error in the level bellow. It means excitatory bottomup inputs to a prediction error neuron with inhibitory synaptic inputs that are driven by top-down predictions. In this hierarchical arrangement (adaptive resonance theory) the problem arises because they ignore uncertainty and decoherence in probabilistic representations. This is because the parameters of hierarchical model determine how expected states (synaptic activity) are mixed to form predictions trade-off. So far, the prediction errors in level above can by top-down links suppress the predictions in level below. This centralized elimination, repression, suppression, resonance, assimilation, dissipation is the core of Darwinian adaptation as a hierarchical conformism. That is why may be Darwinian adaptation only a special case of the broader theory of adaptation.
Another problem with the Darwinian adaptation is given by Quantum Darwinism and phenomenon of decoherence. It means that any coherent superposition of the system’s quantum states is continuously reduced to a mixture. A preferred basis called a “pointer basis” is singled out. An effective counterselection rule has emerged: the decoherence which prevents superpositions of the preferred basis from persisting by a lost of diagonal terms of the density matrix. Reduction to a mixture can’t be interpreted as adaptation, progress, development, forward evolution of the system. It is clearly only a counter selection.
In this case the density matrix p(x, x’)of the particle in the position representations evolves (in this context is different from “evolves” in Mendelian evolution that is described as “change in allele frequency per generation”) according to the master equation
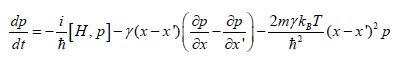
where H is the particle’s Hamiltonian, V(x) is potential, γ is the relaxation rate, kB is the Boltzmann constant, and T is the temperature of the field. This equation naturally separates into three distinct terms, each of them responsible for a different aspect of classical behavior: the first term, the von Neumann equation is derived from the Schrὅdinger equation. The second term causes dissipation: the loss of energy and decrease of the average momentum. The third term is responsible for the fluctuations or random kicks that lead to Brownian motion [14]. They causing the environment-induced decoherence based counter selection.
Negative selection due to decoherence is the essence of environment-induced counter selection of Darwinian adaptation. Under scrutiny of the environment, only pointer states remain unchanged. Other states decohere into mixtures of stable pointer states that can persist, and, in this sense exist: they are counter selected.
These changes are starting at the level of the quantum evolution and then can manifest at the level classical genetic evolution. Only states that produce multiple informational offspring – multiple imprints on the environment – can be found out from the fragments of environment. The origin of the emergent classicality is then not just survival of the fittest states to deposit multiple records – copies of themselves – through environment. Proliferation of records allows information about system to be extracted from many fragments of the environment. Thus, environment acquires redundant records of the system [15].
The inhibitory-proliferative adaptation
On the basis of prior research, these brain regions include the dorsal anterior cingulated cortex (dACC), and the anterior insula. Exposure to an acute episode of social rejection or to rejectionrelated cues (for example the unemployment) has been shown to activate both the dACC and the anterior insula. Greater activity found in the dACC, in turn, has been associated with greater selfreported feelings of social distress [12].
Psychological stress and exposure to the stressor of Darwinian social disruption (SDR), as a cause of carcinogenesis, increase cytokine production by monocytes/macrophages and reduce their sensitivity to corticosterone.
Repeated social defeat during SDR resulted in a significant increase in spleen mass and the number of splenic monocytes/ macrophages and granulocytes. It indicates that repeated social defeat during the SDR stressor enhances innate immunity to E. coli infection and SDR significantly impacts splenic monocytes/ macrophages.
Recent experimental studies have illuminated the mechanistic pathways by which NF- ĸB signaling contributes to the aspects of carcinogenesis. These data showing that chronic inflammation promotes carcinogenesis and that NF-ĸB signaling is at the heart of such inflammation. Despite versatile and occasionally antagonistic interactions, NF-ĸB and STAT3 cooperate to promote the development and progression of colon, gastric and liver cancers.
In our model the values of y1 and y2 are transformed through a nonlinear activation function f (y) before they inhibit each other:
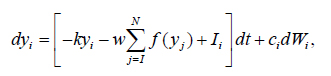
integration starts from yi (0) = 0, an input unit with mean activity Il, and independent white noise fluctuations dWi of amplitude ci. These units also inhibit each other with a connection weight w, k denotes the decay rate of the accumulated activity with leak (entropy), N means the number of alternatives. The inhibition parameter w suppress the self-replicators y3, y4, y5- i.e. counter select the y1, y2.
From a physiological perspective, increased methylation of the Avp enhancer during postnatal life serves to restrain the HPA axis in critical periods when homeostatic thresholds are set, what facilitate adaptation of the endocrine system to environmental stimuli. Phosphorylation of MeCP2 is a conduit of experiencedriven changes in gene expression, serving as an important mediator. Focusing on DNA methylation, provide evidence for postmitotic epigenetic modifications in neuronal function, which facilitate physiological and behavioral adaptations [10].
Conclusions
Intelligent Evolution (iE) means a transition from the hierarchical arrangement to the parallel networking. The hierarchical arrangement leads to stressor social disruption (SDR) which is subsequently contributing to the carcinogenesis.
Despite of Darwin, England and Hawking, for the ongoing type of evolution we propose the label “inhibitory-proliferative adaptation” as a form of Intelligent Evolution (iE) leading to the development of full Artificial Intelligence (AI). Up to date there is no experience for stopping the evolution of the AI prematurely in the half of the way.
“The generation of nonequilibrium structures (such as Bénard vortices or chemical oscillations), where energy is conserved, also corresponds to a free lunch, for the price of nonequilibrium structures is entropy, and not energy.” [7]
The hierarchical arrangement of adaptation with brain at the top may be in contradiction with recent research focus on tumor microenvironment leading to controversies in oncology. On the basis of prior research, these brain regions include the dorsal anterior cingulated cortex (dACC), and the anterior insula. Exposure to an acute episode of social rejection or to rejectionrelated cues (for example the unemployment) has been shown to activate both the dACC and the anterior insula. Greater activity found in the dACC, in turn, has been associated with greater selfreported feelings of social distress [12].
Psychological stress and exposure to the stressor of Darwinian social disruption (SDR), as a cause of carcinogenesis, increase cytokine production by monocytes/macrophages and reduce their sensitivity to corticosterone.
Repeated social defeat during SDR resulted in a significant increase in spleen mass and the number of splenic monocytes/ macrophages and granulocytes. It indicates that repeated social defeat during the SDR stressor enhances innate immunity to E. coli infection and SDR significantly impacts splenic monocytes/ macrophages.
In settings in which hierarchies are strongly enforced, and subordinates have little social support, low dominance rank can lead to chronic stress, immune compromise, reproductive dysregulation and cancer [10, 11, 13].
It is widely accepted that psychological stress affects the immune response, and chronic, repeated exposure to a stressor is immunosuppressive. Ligation of glucocorticoid (GC) receptors on mononuclear cells suppresses the expression of cytokines, chemokines, and adhesion molecules through negative regulation of NF- ĸB activation and function [1].
References
- MT Bailey, H Engler, ND Powell. (2007) Repeated social defeat increases the bactericidal activity of splenic macrophages through a Toll-like receptordependent pathway. Am J Physiol Integr Comp Physiol 293: 1180-1190.
- Mays JW, Bailey MT, Sheridan JF (2010) Influenza Virus-Specific Immunological Memory Is Enhanced by Repeated Social Defeat. J Immunol 184 (4): 2014-2025.
- JL England (2013) Statistical physics of self-replication. J Chem Phys. 139: 121923.
- Engler H, Engler A, Bailey MT, Sheridan JF (2005) Tissue-specific alterations in the glucocorticoid sensitivity of immune cells following repeated social defeat in mice. J Neuroimmunol 163(1-2):110-119.
- Stark JL, Avitsur R, Sheridan JF (2002) Interleukin-6 and the development of social disruption-induced glucocorticoid resistance. J Neuroimmunol 124(1-2): 9-15.
- Avitsur R, Stark JL, Dhabhar FS, Sheridan JF (2002) Social stress alters splenocyte phenotype and function. J Neuroimmunol 132(1-2): 66-71.
- I Prigogine. (1997) The End of Certainty. Time, Chaos, and New Laws of Nature. The Free Press, New York, 175.
- K Friston. (2010) The free-energy principle: a unified brain theory? Nat Rev Neurosci 11(2):127-138.
- DW McShea, RN Brandon (2010) Biology’s First Law. The University of Chicago Press, Chicago.
- C Murgatroyd, AV Patchev, Y Wu. (2009) Dynamic DNA methylation programs persistent adverse effects of early-life stress. Nature Neuroscience 12:1559-1566.
- C Shee, JL Gibson, MC Darrow (2011) Impact of a stress-inducible switch to mutagenic repair of DNA breaks on mutation in Escherichia coli. PNAS 108(33): 13659-13664.
- GM Slavich. (2010) Neural sensitivity to social rejection is associated with inflammatory responses to social stress. PNAS 107(33): 14817-14822.
- J Tung, LB Barreiro, ZP Johnson (2012) Social environment is associated with gene regulatory variation in the rhesus macaque immune system. PNAS 109(17): 6490-6495.
- WH Zurek (1991) Decoherence And The Transition From Quantum To Classical. Physics Today, 44:36-44.
- WH Zurek (2009) Quantum Darwinism. Nature Physics 5:181-188.
- JA Coyne (2009) Why evolution is true. Penguin, New york.
- R Dawkins (2009) The greatest show on earth: The evidence for evolution. Free Press, New York.
- Miller, Stanley L (1953) Production of Amino Acids Under Possible Primitive Earth Conditions. Science 117 (3046): 528-529.
- Zerancic Z, Brenner M (2013) Self-replicating colloidal clusters. PNAS 111(5): 1748-1753.
- Gray M (2010) Irremediable Complexity. Science 330(6006):920-921.
- Crooks G. (1999) Entropy production fluctuation theorem and the nonequilibrium work relation for free energy differences. Physical Review 60:2721.
- Stoltzfus A (1999) On the possibility of constructive neutral evolution. Journal of Molecular Evolution 49(2):169-181.
- Stoltzfus, Arlin (2012) Constructive neutral evolution: exploring evolutionary theory’s curious disconnect. Biology Direct 13(7):35.
- Schrὅdinger E (1967) What is Life ? Cambridge Press, Cambridge.





